I neuroni sono conduttori di elettricità. ☵ Potenziale d'azione dei neuroni: informazioni generali Il marketing è più avanti della scienza
Lascia che ti dica subito che questa nota non ha nulla a che fare con i percettroni, le reti Hopfield o qualsiasi altra rete neurale artificiale. Simuleremo il lavoro di una rete neurale biologica “reale”, “vivente” in cui avvengono i processi di generazione e propagazione degli impulsi nervosi. Nella letteratura in lingua inglese, tali reti, a causa della loro differenza dalle reti neurali artificiali, sono chiamate reti neurali spiking, ma nella letteratura in lingua russa non esiste un nome stabilito. Alcuni le chiamano semplicemente reti neurali, altri le chiamano reti neurali a impulso e altri ancora le chiamano reti di picco.La maggior parte dei lettori avrà probabilmente sentito parlare dei progetti Blue Brain e Human Brain, sponsorizzati dall'Unione Europea, per quest'ultimo progetto il governo dell'UE ha stanziato circa un miliardo di euro, il che indica che c'è un grande interesse in questo settore. Entrambi i progetti sono strettamente correlati e si sovrappongono tra loro, hanno persino lo stesso regista, Henry Markram, il che potrebbe creare confusione su come differiscono l'uno dall'altro. In breve, l’obiettivo finale di entrambi i progetti è sviluppare un modello dell’intero cervello, composto da circa 86 miliardi di neuroni. Il Blue Brain Project è la parte computazionale, e Human Brain è più una parte fondamentale, dove stanno lavorando alla raccolta di dati scientifici sui principi della funzione cerebrale e alla creazione di un modello unificato. Per entrare in contatto con questa scienza e provare a fare noi stessi qualcosa di simile, anche se su scala molto più piccola, è stata scritta questa nota.
Sull'hub sono già stati pubblicati diversi articoli interessanti e informativi sulla neurobiologia, il che è molto piacevole.
1. Neurobiologia e intelligenza artificiale: prima parte - programma didattico.
2. Neurobiologia e intelligenza artificiale: seconda parte - L'intelligenza e la rappresentazione dell'informazione nel cervello.
3. Neuroscienze e intelligenza artificiale: terza parte – Rappresentazione dei dati e memoria
Ma non affrontavano questioni di neuroscienza computazionale, o comunque di neuroscienza computazionale, che include la modellizzazione computerizzata dell’attività elettrica dei neuroni, quindi ho deciso di colmare questa lacuna.
Un po' di biologia
Riso. 1 - Rappresentazione schematica della struttura di un neurone.
Prima di iniziare a creare modelli, dobbiamo familiarizzare con alcune neuroscienze di base. Un tipico neurone è costituito da 3 parti: un corpo (soma), dendriti e un assone. I dendriti ricevono segnali da altri neuroni (questo è l'input del neurone) e l'assone trasmette segnali dal corpo del neurone ad altri neuroni (l'output). Il punto di contatto tra l'assone di un neurone e il dendrite di un altro neurone è chiamato sinapsi. Il segnale ricevuto dai dendriti si riassume nel corpo e se supera una certa soglia si genera un impulso nervoso o, in altre parole, un picco. Il corpo cellulare è circondato da una membrana lipidica, che è un buon isolante. Le composizioni ioniche del citoplasma dei neuroni e del fluido intercellulare differiscono. Nel citoplasma la concentrazione di ioni potassio è maggiore e la concentrazione di sodio e cloro è inferiore, ma nel fluido intercellulare è vero il contrario. Ciò è dovuto al funzionamento delle pompe ioniche, che pompano costantemente determinati tipi di ioni contro un gradiente di concentrazione, consumando energia immagazzinata nelle molecole di adenosina trifosfato (ATP). La più famosa e studiata di queste pompe è la pompa sodio-potassio. Rimuove 3 ioni sodio all'esterno e porta 2 ioni potassio all'interno del neurone. La Figura 2 illustra la composizione ionica di un neurone ed etichetta le pompe ioniche. Grazie al lavoro di queste pompe, nel neurone si forma una differenza di potenziale di equilibrio tra il lato interno della membrana, che è carico negativamente, e quello esterno, carico positivamente. 
Riso. 2 - Composizione ionica del neurone e dell'ambiente
Oltre alle pompe, sulla superficie del neurone ci sono anche canali ionici che, quando cambia il potenziale o quando sono esposti a sostanze chimiche, possono aprirsi o chiudersi, aumentando o diminuendo così le correnti di un certo tipo di ioni. Se il potenziale di membrana supera una certa soglia, i canali del sodio si aprono e poiché all'esterno c'è più sodio, appare una corrente elettrica diretta all'interno del neurone, che aumenta ulteriormente il potenziale di membrana e apre ancora di più i canali del sodio, si verifica un forte aumento del potenziale di membrana . I fisici chiameranno questo feedback positivo. Ma, a partire da un certo valore potenziale superiore al potenziale soglia per l'apertura dei canali del sodio, si aprono anche i canali del potassio, per cui gli ioni potassio iniziano a fluire verso l'esterno, riducendo il potenziale di membrana e riportandolo così al valore di equilibrio. Se l'eccitazione iniziale è inferiore alla soglia per l'apertura dei canali del sodio, il neurone tornerà al suo stato di equilibrio. La cosa interessante è che l’ampiezza dell’impulso generato dipende debolmente dall’ampiezza della corrente eccitante: o l’impulso c’è oppure non c’è, vale la legge del “tutto o niente”.
A proposito, è stato il principio “tutto o niente” a ispirare McCulloch e Pitts a creare modelli di reti neurali artificiali. Ma il campo delle reti neurali artificiali si sta sviluppando a modo suo e il suo obiettivo principale è la soluzione ottimale ai problemi pratici, indipendentemente da come ciò si riferisca ai processi di elaborazione delle informazioni nel cervello vivente. Mentre le reti neurali a spillo sono un modello di come funziona un vero cervello. È possibile assemblare una rete di picchi per il riconoscimento delle immagini visive, ma per l'uso pratico le reti neurali classiche sono più adatte, sono più semplici, vengono calcolate più velocemente su un computer e per loro sono stati inventati molti algoritmi per l'addestramento a compiti pratici specifici.
Il principio "tutto o niente" è chiaramente rappresentato nella Figura 3. In basso c'è la corrente in ingresso diretta all'interno della membrana neuronale, e in alto c'è la differenza di potenziale tra l'interno e l'esterno della membrana. Pertanto, secondo il concetto attualmente dominante nelle reti neurali viventi, l'informazione viene codificata nel momento in cui si verificano gli impulsi o, come direbbero i fisici, attraverso la modulazione di fase. 
Riso. 3 - Generazione di un impulso nervoso. Di seguito è riportata la corrente fornita alla cella in pA e in alto è riportato il potenziale di membrana in mV
È possibile eccitare un neurone, ad esempio, inserendovi un microelettrodo e applicando corrente al suo interno, ma in un cervello vivente l'eccitazione avviene solitamente attraverso l'azione sinaptica. Come già accennato, i neuroni si collegano tra loro utilizzando le sinapsi formate nei punti di contatto dell'assone di un neurone con i dendriti di un altro. Il neurone da cui proviene il segnale è detto presinaptico, mentre quello a cui arriva il segnale è detto postsinaptico. Quando un impulso si verifica su un neurone presinaptico, rilascia neurotrasmettitori nella fessura sinaptica, che aprono canali di sodio sul neurone postsinaptico, e quindi si verifica la catena di eventi sopra descritta, che porta all'eccitazione. Oltre all’eccitazione, i neuroni possono anche inibirsi a vicenda. Se il neurone presinaptico è inibitorio, rilascerà un neurotrasmettitore inibitorio nella fessura sinaptica che apre i canali del cloro e poiché c'è più cloro all'esterno, il cloro fluisce nel neurone a causa del quale aumenta la carica negativa sul lato interno della membrana (non dimenticare che gli ioni cloro a differenza di sodio e potassio sono caricati negativamente), portando il neurone in uno stato ancora più inattivo. In questo stato, il neurone è più difficile da eccitare.
Modello matematico di un neurone
Sulla base dei meccanismi dinamici del neurone sopra descritti, è possibile compilare il suo modello matematico. Al momento sono stati creati vari modelli relativamente semplici, come “Ingregrate and Fire”, in cui un neurone è rappresentato come un condensatore e un resistore, nonché modelli più complessi e biologicamente plausibili, come il modello di Hodgkin-Huxley, che è molto più complesso sia in termini computazionali che in termini di analisi della sua dinamica, ma descrive in modo molto più accurato la dinamica del potenziale di membrana di un neurone. In questo articolo utilizzeremo il modello Izhikevich; esso rappresenta un compromesso tra complessità computazionale e plausibilità biofisica. Nonostante la sua semplicità computazionale, questo modello può riprodurre un gran numero di fenomeni che si verificano nei neuroni reali. Il modello Izhikevich è specificato come un sistema di equazioni differenziali (Figura 4).
Riso. 4 - Modello Izhikevich
Dove a, b, c, d, k, cm vari parametri neuronali. Vmè la differenza di potenziale sui lati interno ed esterno della membrana, e Ehm- variabile ausiliaria. IOè una corrente applicata costante esterna. In questo modello si osservano le proprietà caratteristiche dei neuroni: generazione di un picco in risposta a un singolo impulso di corrente esterna e generazione di una sequenza di picchi con una certa frequenza quando al neurone viene applicata una corrente esterna costante. Isyn- la somma delle correnti sinaptiche di tutti i neuroni a cui questo neurone è connesso.
Se viene generato un picco su un neurone presinaptico, si verifica un salto di corrente sinaptica sul neurone postsinaptico, che decade esponenzialmente con un tempo caratteristico.
Passiamo alla codifica
Quindi arriviamo alla parte divertente. È ora di codificare un pezzo virtuale di tessuto neurale sul computer. Per fare ciò, risolveremo numericamente un sistema di equazioni differenziali che definiscono la dinamica del potenziale di membrana di un neurone. Per l'integrazione utilizzeremo il metodo di Eulero. Codificheremo in C++, disegneremo utilizzando script scritti in Python utilizzando la libreria Matplolib, ma chi non ha Python può disegnare utilizzando Excel.Avremo bisogno di array bidimensionali Vms, ehm dimensioni Tsim*Nneur immagazzinare i potenziali di membrana e le variabili ausiliarie di ciascun neurone, in ogni momento, Tsim questo è il tempo di simulazione in conteggi e Nneur numero di neuroni nella rete.
Memorizzeremo le connessioni sotto forma di due array pre_con E post_con dimensioni Non c'è ne, dove gli indici sono i numeri di connessioni e i valori sono gli indici dei neuroni presinaptici e postsinaptici. Non c'è ne- numero di connessioni.
Abbiamo anche bisogno di un array per rappresentare una variabile che moduli la corrente postsinaptica che decade esponenzialmente di ciascuna sinapsi, per questo creiamo un array sì dimensioni Ncon*Tsim.
Galleggiante costante h = 0,5f; // passo temporale di integrazione in ms const int Tsim = 1000/.5f; // tempo di simulazione in campioni discreti const int Nexc = 100; // Numero di neuroni eccitatori const int Ninh = 25; // Numero di neuroni inibitori const int Nneur = Nexc + Ninh; const int Ncon = Nneur*Nneur*0.1f; // Numero di connessioni, 0,1 è la probabilità di comunicazione tra 2 neuroni casuali float Vms; // i potenziali di membrana galleggiano Ums; // variabili ausiliarie del modello Izhikevich float Iex; // corrente continua esterna applicata al neurone float Isyn; // corrente sinaptica per neurone int pre_conns; // indici dei neuroni presinaptici int post_conns; // gli indici dei neuroni postsinaptici fluttuano i pesi; // i pesi delle connessioni float y; // corrente sinaptica modulante variabile in base ai picchi nella presinapsi float psc_excxpire_time = 4.0f; // tempo caratteristico di decadimento della corrente postsinaptica, ms float minWeight = 50.0f; // pesi, dimensione pcA float maxWeight = 100.0f; // I parametri del neurone float Iex_max = 40.0f; // la corrente massima applicata al neurone è 50 pA float a = 0,02f; float b = 0,5f; float c = -40.0f; // il valore del potenziale di membrana a cui viene reimpostato dopo un picco float d = 100.0f; galleggiante k = 0,5f; float Vr = -60.0f; float Vt = -45.0f; float Vpicco = 35.0f; // il valore massimo del potenziale di membrana al quale avviene il ripristino al valore con float V0 = -60.0f; // valore iniziale per il potenziale di membrana float U0 = 0.0f; // valore iniziale per la variabile ausiliaria float Cm = 50.0f; // capacità elettrica di un neurone, dimensione pcF
Come già accennato, l'informazione viene codificata nei momenti in cui si verifica l'impulso, quindi creiamo array per memorizzare i tempi del loro verificarsi e gli indici dei neuroni in cui si sono verificati. Quindi possono essere scritti in un file a scopo di visualizzazione.
Spike_times in virgola mobile; // tempi in cui si sono verificati i picchi int spike_neurons; // indici dei neuroni su cui si verificano gli picchi int spike_num = 0; // numero di picco
Spargiamo casualmente le connessioni e impostiamo i pesi.
Void init_connections())( for (int con_idx = 0; con_idx< Ncon;){
// случайно выбираем постсипантические и пресинаптические нейроны
pre_conns = rand() % Nneur;
post_conns = rand() % Nneur;
weights = (rand() % ((int)(maxWeight - minWeight)*10))/10.0f + minWeight;
if (pre_conns >= Nexc)( // se il neurone presinaptico è inibitorio, allora il peso della connessione ha un segno menoweights = -weights; ) con_idx++; ) )
Impostazione delle condizioni iniziali per i neuroni e assegnazione casuale della corrente applicata esterna. Quei neuroni per i quali la corrente esterna supera la soglia di generazione dei picchi genereranno picchi a frequenza costante.
Void init_neurons())( for (int neur_idx = 0; neur_idx< Nneur; neur_idx++){
// случайно разбрасываем приложенные токи
Iex = (rand() % (int) (Iex_max*10))/10.0f;
Isyn = 0.0f;
Vms = V0;
Ums = U0;
}
}
La parte principale del programma con l'integrazione del modello Izhikevich.
Float izhik_Vm(int neurone, int ora)( return (k*(Vms - Vr)*(Vms - Vt) - Ums + Iex + Isyn)/Cm; ) float izhik_Um(int neurone, int ora)( return a*( b*(Vms - Vr) - Ums); ) int main())( init_connections(); init_neurons(); float scadere_coeff = exp(-h/psc_excxpire_time); // per la corrente che decade esponenzialmente per (int t = 1; T< Tsim; t++){
// проходим по всем нейронам
for (int neur = 0; neur < Nneur; neur++){
Vms[t] = Vms + h*izhik_Vm(neur, t-1);
Ums[t] = Ums + h*izhik_Um(neur, t-1);
Isyn = 0.0f;
if (Vms >Vpeak)( Vms[t] = c; Ums[t] = Ums + d; picco_times = t*h; picco_neurons = neur; picco_num++; ) ) // passa attraverso tutte le connessioni per (int con = 0; con< Ncon; con++){
y[t] = y*expire_coeff;
if (Vms] >Vpeak)( y[t] = 1.0f; ) Isyn] += y[t]*pesi; ) ) salva2file(); restituire 0; )
È possibile scaricare il testo completo del codice

Per trasportare un segnale dalla cellula precedente a quella successiva, il neurone genera segnali elettrici al suo interno. I movimenti degli occhi durante la lettura di questo paragrafo, la sensazione di una sedia morbida sotto il sedere, la percezione della musica dalle cuffie e molto altro si basano sul fatto che dentro di te passano centinaia di miliardi di segnali elettrici. Un segnale di questo tipo può avere origine nel midollo spinale e viaggiare fino alla punta del piede lungo un lungo assone. Oppure può percorrere una distanza trascurabile nelle profondità del cervello, limitandosi ai confini di un interneurone con processi brevi. Qualsiasi neurone che riceve un segnale lo guida attraverso il suo corpo e i suoi processi, e questo segnale è di natura elettrica.
Già nel 1859 gli scienziati furono in grado di misurare la velocità con cui venivano trasmessi questi segnali elettrici. Si è scoperto che l'elettricità trasmessa lungo un assone vivente è fondamentalmente diversa dalla corrente elettrica nei metalli. Un segnale elettrico viene trasmesso attraverso un filo metallico ad una velocità prossima a quella della luce (300.000 chilometri al secondo), perché nel metallo ci sono molti elettroni liberi. Tuttavia, nonostante questa velocità, il segnale si indebolisce notevolmente quando si percorrono lunghe distanze. Se i segnali venissero trasmessi lungo gli assoni nello stesso modo in cui vengono trasmessi nei metalli, l'impulso nervoso proveniente dalla terminazione nervosa nella pelle dell'alluce verrebbe completamente attenuato prima di raggiungere il cervello - anche la resistenza elettrica della materia organica lo è alto e il segnale è troppo debole.
La ricerca ha dimostrato che l'elettricità viene trasmessa attraverso gli assoni molto più lentamente che attraverso i fili e che questa trasmissione si basa su un meccanismo precedentemente sconosciuto, in seguito al quale il segnale viaggia ad una velocità di circa 30 metri al secondo. I segnali elettrici che viaggiano lungo i nervi, a differenza dei segnali che viaggiano lungo i fili, non si indeboliscono durante il viaggio. La ragione di ciò è che le terminazioni nervose non trasmettono passivamente il segnale attraverso se stesse, ma permettono semplicemente alle particelle cariche che contengono di trasmetterlo l'una all'altra. In ogni momento sono emettitori attivi di questo segnale, che lo trasmettono, e una descrizione dettagliata di questo meccanismo richiederà un capitolo a parte. Pertanto, sacrificando l'elevata velocità degli impulsi nervosi, attraverso la trasmissione attiva del segnale, il neurone riceve la garanzia che il segnale proveniente dall'alluce raggiungerà il midollo spinale senza indebolirsi affatto.

Osservare il passaggio di un'onda di eccitazione elettrica, o potenziale d'azione (potenziale d'azione ['ækʃən pə'tenʃəl]), in una cellula vivente è sufficiente un semplice dispositivo: un'estremità di un sottile filo metallico è posizionata sulla superficie esterna dell'assone di un neurone sensoriale nella pelle, e l'altra è collegata a un registratore che traccia una linea verso l'alto quando il segnale è rafforzato e verso il basso quando il segnale è indebolito. Ogni tocco sulla pelle produce uno o più potenziali d'azione. Quando si verifica ogni potenziale, il registratore disegna un picco lungo e stretto.
Il potenziale d'azione di un neurone sensoriale dura solo circa 0,001 secondi e comprende due fasi: un rapido aumento che raggiunge il picco, e poi un decadimento quasi altrettanto rapido dell'eccitazione, che porta alla posizione iniziale. E poi il registratore riporta un fatto inaspettato: tutti i potenziali d'azione che sorgono nella stessa cellula nervosa sono più o meno gli stessi. Lo si può vedere nell'immagine a sinistra: tutti i picchi disegnati dal registratore hanno all'incirca la stessa forma e ampiezza, indipendentemente da quanto forte e prolungato sia stato il contatto sulla pelle che li ha provocati. Una carezza delicata o un pizzicotto palpabile saranno trasmessi da potenziali d'azione della stessa grandezza. Il potenziale d'azione è un segnale costante che obbedisce al principio “tutto o niente”: dopo che lo stimolo supera un certo valore di soglia, appare sempre approssimativamente lo stesso segnale, né più né meno del solito. E se lo stimolo è inferiore al valore di soglia, il segnale non verrà trasmesso affatto: ad esempio, puoi toccare la pelle con la punta di una penna così facilmente che questo tocco non si sentirà.
Il principio tutto o niente nella generazione dei potenziali d'azione solleva nuove domande. Come fa un neurone sensoriale a comunicare la forza di uno stimolo: pressione forte o debole, luce intensa o fioca? Come comunica la durata dello stimolo? Infine, come fanno i neuroni a distinguere un tipo di informazione sensoriale da un altro? Ad esempio, come distinguono il tatto dal dolore, dalla luce, dall'olfatto o dal suono? E come differenziano le informazioni sensoriali per la percezione dalle informazioni motorie per l'azione?

L'evoluzione ha risolto il problema di come comunicare la forza di uno stimolo utilizzando lo stesso tipo di segnali della stessa grandezza: questa forza è determinata da frequenza(frequenza [‘friːkwənsɪ]), con la quale vengono emessi i potenziali d’azione. Uno stimolo debole, come un leggero tocco sul braccio, provoca l’emissione di soli due o tre potenziali d’azione al secondo, mentre una forte pressione, come un pizzicotto o un colpo al gomito, può causare un’esplosione di centinaia di potenziali d’azione. al secondo. In questo caso, la durata della sensazione è determinata dalla durata della comparsa dei potenziali d'azione.
I neuroni utilizzano codici elettrici diversi per comunicare al cervello che trasportano informazioni su stimoli diversi, come dolore, luce o suono? Si è scoperto che no! Sorprendentemente, c'è pochissima differenza tra i potenziali d'azione generati dai neuroni di diversi sistemi sensoriali (ad esempio, visivo o tattile)! Pertanto, la natura e la natura della sensazione non dipendono dalle differenze nei potenziali d'azione (il che apre una prospettiva piuttosto entusiasmante per pensare al tema della “matrice” del film con lo stesso nome). Il neurone che trasmette l'informazione uditiva è costruito esattamente come il neurone del circuito visivo e conducono gli stessi potenziali d'azione nello stesso modo. Senza sapere a quale circuito nervoso appartiene un particolare neurone, è impossibile determinare quali informazioni trasporta solo analizzando il suo funzionamento.
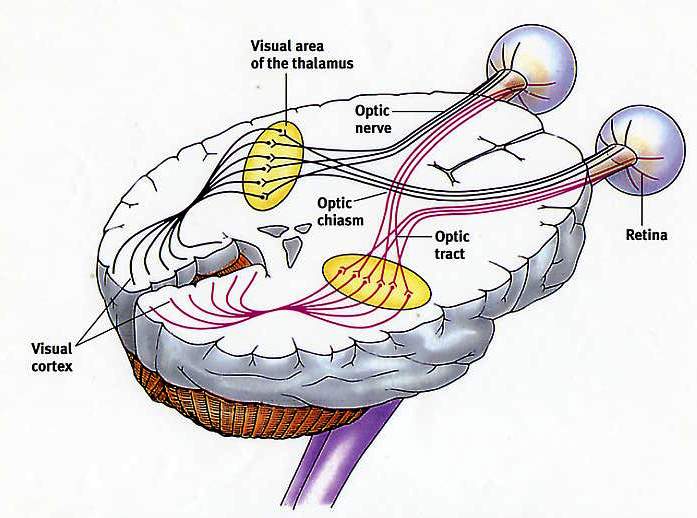
La natura dell'informazione trasmessa dipende principalmente dal tipo di fibre nervose eccitate e dagli specifici sistemi cerebrali a cui queste fibre sono collegate. Le sensazioni di ciascun tipo vengono trasmesse lungo i propri percorsi e il tipo di informazione trasmessa da un neurone dipende dal percorso che include questo neurone. In qualsiasi percorso sensoriale, le informazioni vengono trasferite dal primo neurone sensoriale (un recettore che risponde a uno stimolo esterno come il tatto, l'olfatto o la luce) ai neuroni specializzati nel midollo spinale o nel cervello. Pertanto, l'informazione visiva differisce da quella uditiva solo perché viene trasmessa lungo altri percorsi, partendo dalla retina e terminando nella parte del cervello responsabile della percezione visiva.
Anche i segnali inviati dai motoneuroni nel cervello ai muscoli sono quasi identici a quelli inviati lungo i neuroni sensoriali dalla pelle al cervello. Obbediscono allo stesso principio "tutto o niente", trasmettono anche l'intensità del segnale utilizzando la frequenza dei potenziali d'azione e il risultato del segnale dipende solo dal circuito neurale in cui è incluso questo neurone. Pertanto, una rapida serie di potenziali d'azione lungo un percorso specifico provoca il movimento delle dita e non, ad esempio, la percezione di luci multicolori, solo perché questo percorso è collegato ai muscoli delle mani e non alla retina degli occhi.

L'universalità dei potenziali d'azione non si limita alla somiglianza della loro manifestazione in diversi neuroni situati nello stesso organismo. Sono così identici nei diversi animali che anche un ricercatore esperto non è in grado di distinguere con precisione tra la registrazione del potenziale d'azione della fibra nervosa di una balena, di un topo, di una scimmia o del suo supervisore scientifico. Tuttavia, i potenziali d’azione nelle diverse cellule non sono identici: ci sono ancora piccole differenze nella loro ampiezza e durata, e l’affermazione “tutti i potenziali d’azione sono uguali” è tanto imprecisa quanto “tutte le buganvillee sono uguali”.
Quindi, ogni neurone trasmette un segnale attraverso il suo corpo e lo elabora allo stesso modo. Tutta la varietà di informazioni che riceviamo dai neuroni sensoriali, tutti i movimenti che il nostro corpo può compiere, sono il risultato della trasmissione di un unico tipo di segnali all'interno dei neuroni. Resta solo una piccola cosa: capire che tipo di segnale è e come viene trasmesso.

Separiamo abitualmente tutto ciò che consideriamo natura vivente, compresi noi stessi, dalle cose “non viventi”, compresi i metalli e la corrente elettrica trasmessa attraverso di essi. È ancora più sorprendente rendersi conto che i metalli non sono solo presenti nel nostro corpo, ma sono necessari, senza di essi il corpo non può esistere. La corrente elettrica non è un fenomeno occasionale, ma si verifica continuamente in centinaia di miliardi di neuroni che permeano tutto il nostro corpo con i loro processi. In questo momento puoi avvertire diversi segni della sua presenza: la tua consapevolezza di questo testo è il risultato di innumerevoli trasmissioni di corrente elettrica. La sensazione di fame e di piacere derivante dall'odore del cibo cucinato, la percezione stessa di questo odore, il tocco del vento che vola attraverso la finestra sulla tua pelle... La lista è infinita. E il desiderio di capire come tutto ciò avvenga consiste anche negli impulsi elettrici che nascono nei neuroni.
Poiché lo scopo di questo capitolo è riportare solo le informazioni più generali sul passaggio di un impulso nervoso, è necessario considerare anche l'ambiente in cui esso nasce, quelle condizioni nella cellula che ne rendono possibile la comparsa e la trasmissione. Vale quindi la pena iniziare studiando il trampolino di lancio su cui si svilupperanno gli eventi, ovvero dal neurone in entrata a riposo (stato dormiente ['dɔːmənt steɪt]).

Già a metà del secolo scorso, gli scienziati trovarono un modo per determinare in quale parte di un neurone esiste una carica elettrica. Per questo usano voltmetro (voltmetro ['vəultˌmiːtə]) (un dispositivo per misurare la tensione del campo elettrico) con due elettrodi. Un elettrodo viene posizionato all'interno del neurone, posizionandolo vicino alla membrana cellulare, mentre il secondo elettrodo viene posizionato nell'ambiente circostante il neurone, dall'altra parte della stessa membrana. Il voltmetro mostra che ci sono cariche elettriche, negativo all'interno della cellula e positivo all'esterno. L'esistenza di tali cariche elettriche opposte polari su entrambi i lati della membrana crea un campo elettrico, una caratteristica importante del quale è potenziale. Il potenziale, in termini semplici, è la capacità di compiere un lavoro, come il lavoro di trascinare una particella carica da un luogo all'altro. Più cariche negative si accumulano su un lato e più cariche positive si accumulano sull'altro lato della membrana, più forte è il campo elettrico che creano e maggiore è la forza che riescono a trascinare avanti e indietro le particelle cariche. Si chiama la differenza tra le cariche elettriche esterne e interne potenziale di membrana (potenziale di membrana ['membreɪn pə'tenʃəl]) pace. Per un neurone è di circa 70 mV (millivolt), cioè 70 millesimi di volt o sette centesimi di volt. Per fare un confronto, la differenza di potenziale in una batteria AA è di 1,5 volt, ovvero 20 volte maggiore. Cioè, il potenziale di membrana a riposo di un neurone è solo 20 volte più debole di quello tra i terminali di una batteria AA - piuttosto grande, a quanto pare. Il potenziale elettrico esiste solo sulla membrana e in altre parti del neurone è elettricamente neutro.
Per essere più precisi, il potenziale di membrana a riposo di un neurone è -70 mV (meno settanta millivolt). Il segno meno significa solo che la carica negativa si trova all'interno della cellula, e non all'esterno, e quindi il campo elettrico creato è in grado di trascinare ioni con carica positiva attraverso la membrana nella cellula.
Attori nella creazione del potenziale di membrana a riposo:

1 . IN membrana cellulare neurone ci sono canali attraverso i quali possono viaggiare gli ioni che trasportano una carica elettrica. Inoltre, la membrana non è solo un “divisorio” passivo tra l’ambiente interno del neurone e il fluido intercellulare che lo circonda: speciali proteine incorporate nella carne della membrana aprono e chiudono questi canali, e quindi la membrana controlla il passaggio degli ioni - atomi dotati di carica elettrica. Accumulando ioni caricati negativamente all'interno della cellula, il neurone aumenta il numero di cariche negative all'interno, provocando così un aumento delle cariche positive all'esterno e quindi aumentando il potenziale elettrico. Poiché un protone ha una carica positiva e un elettrone una carica negativa, un eccesso di protoni produce uno ione carico positivamente, mentre un eccesso di elettroni produce uno ione carico negativamente. Se desideri informazioni più dettagliate su atomi e ioni, puoi tornare a. È importante capire che il potenziale di membrana esiste proprio al confine della membrana cellulare e che i fluidi generalmente all'interno e all'esterno del neurone rimangono elettricamente neutri. Gli ioni per i quali la membrana è permeabile rimangono vicini ad essa, poiché le cariche positive e negative si attraggono reciprocamente. Di conseguenza, all'esterno della membrana si forma uno strato di ioni positivi "seduti" su di essa e all'interno ioni negativi. Pertanto, la membrana svolge il ruolo di una capacità elettrica che separa le cariche, all'interno della quale è presente un campo elettrico. La membrana è quindi un condensatore naturale.
2 . proteine caricate negativamente, situato all'interno del neurone vicino alla superficie interna della membrana. La carica delle proteine rimane sempre la stessa e costituisce solo una parte della carica totale della superficie interna della membrana. A differenza degli ioni, le proteine non possono uscire o entrare in una cellula perché sono troppo grandi per questo. La carica totale cambia a seconda del numero di ioni carichi positivamente situati vicino alla membrana, la cui concentrazione può cambiare a causa della loro transizione dalla cellula all'esterno e dall'esterno all'interno.
3 . Gli ioni potassio (K+) caricati positivamente possono muoversi liberamente tra l'ambiente interno ed esterno quando il neurone è a riposo. Si muovono costantemente aperti canali di flusso del potassio (passaggio del flusso del potassio), attraverso il quale possono passare solo gli ioni K+ e nient'altro. I canali di flusso sono quelli che non hanno una porta, il che significa che sono aperti in qualsiasi stato del neurone. Ci sono molti più ioni potassio all'interno della cellula che all'esterno. Ciò avviene a causa del funzionamento costante della pompa sodio-potassio (ne parleremo più avanti), quindi, nello stato di riposo del neurone, gli ioni K + iniziano a spostarsi nell'ambiente esterno, poiché la concentrazione della stessa sostanza tende livellarsi nel sistema generale. Se versiamo una sostanza in una pozza d'acqua in un angolo, la sua concentrazione in quest'angolo sarà molto alta e in altre parti della piscina sarà zero o molto piccola. Tuttavia, dopo un po' di tempo scopriremo che la concentrazione di questa sostanza si è stabilizzata in tutto il bacino a causa del moto browniano. In questo caso si parla di “pressione parziale” di una sostanza, sia essa un liquido o un gas. Se l'alcol viene versato in un angolo della piscina, ci sarà una grande differenza nella concentrazione di alcol tra quell'angolo e il resto della piscina. Si creerà una pressione parziale delle molecole di alcol, che si distribuiranno gradualmente in modo uniforme in tutta la piscina in modo che la pressione parziale scomparirà, poiché la concentrazione delle molecole di alcol sarà uniforme ovunque. Pertanto, gli ioni K + portano via la carica positiva dal neurone, spostandosi verso l'esterno a causa della pressione parziale, che è più forte della forza di attrazione delle proteine caricate negativamente, se la differenza nella concentrazione di ioni all'interno e all'esterno della cellula è sufficientemente grande. Poiché le proteine caricate negativamente rimangono all'interno, si forma una carica negativa sul lato interno della membrana. Per una chiara comprensione del funzionamento dei meccanismi cellulari, è importante ricordare che, nonostante il flusso costante di ioni di potassio dalla cellula, ce ne sono sempre di più all'interno del neurone che all'esterno.
4 . Gli ioni sodio caricati positivamente (Na+) si trovano all'esterno della membrana e lì creano una carica positiva. Durante la fase di riposo del neurone, i canali del sodio della cellula Chiuso e Na + non può passare all'interno e la loro concentrazione all'esterno aumenta a causa del lavoro della pompa sodio-potassio, che li rimuove dal neurone.
5 . il ruolo di carica negativa ioni cloro (Cl -) e carico positivamente ioni calcio (Ca 2+) per creare una membrana il potenziale è piccolo, quindi per ora il loro comportamento rimarrà dietro le quinte.
Formazione del potenziale di membrana a riposo avviene in due fasi:
Fase I. viene creata una piccola differenza di potenziale (-10 mV). pompa sodio-potassio.

A differenza di altri canali di membrana, il canale sodio-potassio è in grado di consentire il passaggio sia degli ioni sodio che di potassio. Inoltre, Na+ può attraversarla solo dalla cellula verso l'esterno, e K+ dall'esterno verso l'interno. Un ciclo di funzionamento di questo canale comprende 4 passaggi:
1 . La "porta" del canale sodio-potassio è aperta solo sul lato interno della membrana e lì entrano 3 Na +

2 . la presenza di Na+ all'interno del canale lo influenza in modo tale da poter distruggere parzialmente una molecola ATP(ATP) ( adenosina trifosfato), (adenosina trifosfato [ə'dɛnəsiːn trai'fɔsfeɪt]) che è la “batteria” della cellula, immagazzinando energia e rilasciandola quando necessario. Con tale distruzione parziale, che consiste nel distacco di un gruppo fosfato PO 4 3− dall'estremità della molecola, viene rilasciata energia, che viene spesa proprio per il trasferimento di Na + nello spazio esterno.
3 . quando il canale si apre per far fuoriuscire il Na+, rimane aperto e vi entrano due ioni K+, attratti dalle cariche negative delle proteine dal suo interno. Il fatto che un canale contenente tre ioni sodio possa accogliere solo due ioni potassio è abbastanza logico: l'atomo di potassio ha un diametro maggiore.
4 . la presenza di ioni potassio ora, a sua volta, influenza il canale in modo che il “cancello” esterno si chiuda, quello interno si apra e il K + entri nell'ambiente interno del neurone.
Ecco come funziona la pompa sodio-potassio, che “scambia” tre ioni sodio con due ioni potassio. Poiché la carica elettrica di Na + e K + è la stessa, risulta che tre cariche positive vengono rimosse dalla cella e solo due entrano all'interno. A causa di ciò, la carica positiva interna della membrana cellulare diminuisce e quella esterna aumenta. Inoltre, si crea una differenza nella concentrazione di Na+ e K+ sui diversi lati della membrana:
=) ci sono molti ioni sodio all'esterno della cellula e pochi all'interno. Allo stesso tempo, i canali del sodio sono chiusi e il Na + non può rientrare nella cellula e non si allontana dalla membrana, poiché è attratto dalla carica negativa esistente all'interno della membrana.
=) ci sono molti ioni potassio all'interno della cellula, ma ce ne sono pochi all'esterno, e questo porta alla fuoriuscita di K + dalla cellula attraverso i canali del potassio che sono aperti durante la fase di riposo del neurone.

Fase II la formazione del potenziale di membrana a riposo si basa proprio su questo deflusso di ioni potassio dal neurone. La figura a sinistra mostra la composizione ionica della membrana all'inizio del secondo stadio della formazione del potenziale di riposo: molto K + e proteine caricate negativamente (designate A 4-) all'interno e Na + aggrappato al membrana all'esterno. Muovendosi nell'ambiente esterno, gli ioni potassio portano via le loro cariche positive dalla cellula, mentre la carica totale della membrana interna diminuisce. Proprio come gli ioni positivi di sodio, gli ioni di potassio fuoriusciti dalla cellula rimangono all'esterno della membrana, attratti dalla carica negativa interna, e la carica positiva esterna della membrana è la somma delle cariche Na + e K +. Nonostante il flusso attraverso i canali di flusso, all'interno della cellula si trovano sempre più ioni potassio che all'esterno.
Sorge la domanda: perché gli ioni potassio non continuano a fuoriuscire finché la loro quantità all'interno e all'esterno della cellula non diventa uguale, cioè finché non scompare la pressione parziale creata da questi ioni? La ragione di ciò è che quando K+ lascia la cellula, si verifica un aumento di carica positiva all'esterno e un eccesso di carica negativa all'interno. Ciò riduce il desiderio degli ioni potassio di lasciare la cellula, perché la carica positiva esterna li respinge e la carica negativa interna li attrae. Pertanto, dopo un po ', K + smette di fuoriuscire, nonostante nell'ambiente esterno la loro concentrazione sia inferiore rispetto a quella interna: l'influenza delle cariche sui lati opposti della membrana supera la forza della pressione parziale, cioè supera il desiderio del K+ di essere distribuito uniformemente nel liquido all'interno e all'esterno del neurone. Nel momento in cui viene raggiunto questo equilibrio, il potenziale di membrana del neurone si ferma a circa -70 mV.
Una volta che un neurone ha raggiunto il potenziale di membrana a riposo, è pronto per avviare e condurre un potenziale d'azione, che sarà discusso nel prossimo capitolo citologico.
Quindi, per riassumere: la distribuzione non uniforme degli ioni potassio e sodio su entrambi i lati della membrana è causata dall'azione di due forze concorrenti: a) la forza di attrazione e repulsione elettrica, e b) la forza di pressione parziale derivante dalla differenza di concentrazioni. Il lavoro di queste due forze concorrenti avviene in presenza di canali del sodio, del potassio e del sodio-potassio diversamente strutturati, che agiscono come regolatori dell'azione di queste forze. Il canale del potassio è un canale di flusso, cioè è sempre aperto quando il neurone è a riposo, in modo che gli ioni K+ possano facilmente muoversi avanti e indietro sotto l'influenza delle forze elettriche di repulsione/attrazione e sotto l'influenza delle forze causate da pressione parziale, cioè la differenza nella concentrazione di questi ioni. I canali del sodio sono sempre chiusi quando il neurone è a riposo, quindi gli ioni Na+ non possono attraversarli. E infine il canale sodio-potassio, progettato in modo da funzionare come una pompa, che ad ogni ciclo espelle fuori tre ioni sodio e spinge all'interno due ioni potassio.
L'intera struttura garantisce la presenza del potenziale di membrana a riposo del neurone: cioè uno stato in cui si ottengono due cose:
a) all'interno c'è una carica negativa e all'esterno una carica positiva.
b) all'interno ci sono molti ioni K +, aggrappati alle parti caricate negativamente delle proteine, e quindi si verifica la pressione parziale del potassio - il desiderio degli ioni di potassio di uscire per equalizzare la concentrazione.
c) all'esterno ci sono molti ioni Na+, che in parte formano coppie con ioni Cl-. E così nasce la pressione parziale del sodio: il desiderio degli ioni sodio di entrare nella cellula per equalizzare la concentrazione.
Come risultato del funzionamento della pompa potassio-sodio, otteniamo tre forze esistenti sulla membrana: la forza del campo elettrico e la forza di due pressioni parziali. Queste forze iniziano a funzionare quando il neurone lascia lo stato di riposo.
Lungo i nervi (singole fibre nervose), i segnali si propagano sotto forma di potenziali d'azione e potenziali elettrotonici, ma su distanze massime diverse. La capacità degli assoni e dei dendriti, nonché delle membrane delle cellule muscolari, di condurre segnali elettrici è caratterizzata dalla loro proprietà del cavo.
Le proprietà dei cavi dei conduttori nervosi sono molto importanti per la propagazione dei segnali nel sistema nervoso. Determinano la generazione di potenziali d'azione nelle terminazioni nervose sensoriali, o recettori, sotto l'azione degli stimoli, la conduzione dei segnali lungo gli assoni e la somma dei segnali da parte della membrana del soma neuronale.
Le moderne teorie sulla conduzione via cavo dell'eccitazione si basano sull'ipotesi di Hermann sull'esistenza di correnti circolari (correnti di Hermann) che fluiscono dalle aree non eccitate della membrana a quelle eccitate quando un impulso si propaga lungo l'assone.
Dalla citologia è noto che ogni cellula eccitabile è delimitata da una membrana plasmatica, alla quale sono adiacenti le membrane che circondano la cellula. Molto spesso, le fibre nervose sono circondate da una guaina mielinica, formata da una cellula gliale nel sistema nervoso centrale o da una guaina formata da una cellula di Schwann nella periferia. Nei siti in cui si ramifica l'assone, o nella sua parte iniziale o terminale, la guaina mielinica diventa più sottile. La membrana stessa è costituita da lipidi e proteine. Tutto ciò determina l'elevata resistenza elettrica della membrana cellulare e la sua elevata capacità elettrica distribuita. Queste caratteristiche determinano le proprietà conduttive della fibra nervosa.
I modelli di base della propagazione dei potenziali, principalmente elettrotonici, lungo le fibre nervose sono stati ottenuti in esperimenti su grandi assoni di calamari. Si è scoperto che quando si applica uno stimolo rettangolare in un certo punto della fibra, il segnale viene registrato con una distorsione man mano che si allontana dal sito di stimolazione. Da un lato si verifica un cambiamento nella forma dei suoi bordi di entrata e di uscita (un ritardo nel raggiungimento del valore massimo) e una diminuzione della sua ampiezza. La prima di queste quantità è determinata dalla costante di tempo, la seconda dalla costante di lunghezza. È noto dalla radiofisica che la costante di tempo di un circuito elettrico avente capacità (C) e resistenza R è determinata dalla formula
τ = RC
e si misura in secondi.
Da cosa dipende la resistenza di una membrana cellulare? Nella cellula ci sono tre percorsi lungo i quali la corrente può fluire in direzione longitudinale lungo l'assone
a) assoplasma
b) liquido extracellulare
c) la membrana stessa
Il liquido extracellulare è un elettrolita, la sua resistenza è bassa. La resistività di una membrana spessa 100 angstrom si avvicina a 1000-5000 ohm × cm, molto grande. La resistività dell'assoplasma è bassa, 200 ohm × cm La capacità delle membrane delle cellule eccitabili C è vicina a 1 μF/cm 2, ma non supera 7 μF/cm 2. Così, τ forse 0,1-7ms. La costante di tempo determina la velocità di ritardo nello sviluppo del potenziale al valore massimo e la velocità di ritardo della sua attenuazione al valore di fondo.
Pendenza L'aumento di potenziale (carica del condensatore a membrana) è determinato dalla legge esponenziale:
V/V 0 =(1-e –t/τ)
L'entità del potenziale V t al tempo t è inferiore al potenziale iniziale V 0 della quantità determinata dall'espressione (1-e – t / τ).
Accettiamo allora t=τ
V t /V 0 =(1-e –1)= 1-1/e=1-1/2,7=0,63
O il 63% dell'originale.
La scarica del condensatore a membrana è descritta anche dalla formula esponenziale:
V t /V 0 =e –t/τ
Prendiamo t=τ, quindi V t /V 0 =e –1 =1/2,7=0,37 o il 37% del massimo dopo timet.
Se attraverso la membrana cellulare scorre una corrente capacitiva di origine elettrotonica, per ogni periodo di tempo pari a τ, la costante di tempo, il segnale dell'elettrotone aumenta del 63% del precedente quando il segnale aumenta, oppure diminuisce al 37% del precedente valore quando diminuisce.
Il meccanismo ionico di questo fenomeno può essere semplicisticamente descritto come segue. Quando le cariche positive vengono introdotte nella cellula (depolarizzazione), gli ioni K+ iniziano a muoversi verso la membrana, che ha una capacità che consente a queste cariche di accumularsi, ma ci sono canali di perdita aperti che consentono agli ioni di passare attraverso e smorzare l'accumulo di carica. Perché si verifichi un vero spostamento di carica, è necessario che passi del tempo. È inoltre necessario del tempo per ripristinare la carica originale quando il condensatore a membrana è scarico. Questo è quello che è τ.
Fino a che punto il potenziale elettrotonico può diffondersi lungo la membrana della fibra nervosa?
La propagazione passiva del segnale dell'elettrotone è determinata dall'equazione U x =U 0 ×e - x /λ, nella quale vediamo ancora una dipendenza esponenziale.
Non è difficile trasformare la formula per il caso x = λ e assicurarsi che il potenziale elettrotonico U x in un punto situato a una distanza x da quello originale sarà inferiore all'originale U 0 di un fattore (fino a 37 % di U 0, cioè ½,7), se questo punto è uguale lunghezza costante λ.
Costante di lunghezza λ , ovvero la costante di polarizzazione spaziale dipende dalla resistenza della membrana r m, dalla resistenza dell'ambiente esterno r o e dalla resistenza assoplasmica r i.
Maggiore è la resistenza della membrana, minore è la resistenza del mezzo, maggiore è la distanza di trasmissione del potenziale elettrotonico. Viene chiamata la diminuzione della grandezza dell'elettrone in base alla distanza decremento .
Si è scoperto che la costante di lunghezza è influenzata dal diametro del conduttore, poiché la resistenza dell'assoplasma dipende da esso. Pertanto, le fibre nervose spesse hanno una distanza λ maggiore sulla quale l'elettrotone può propagarsi.
La propagazione di un elettrotone, principalmente un catelettrotone, è un importante fenomeno fisiologico. Nelle cellule che non generano PD (glia, epitelio, fibre muscolari toniche), a causa della conduzione dell'elettrotone, si realizza una relazione funzionale tra le cellule. Nell'albero dendritico dei neuroni, ad esempio nella corteccia cerebrale, i segnali sotto forma di elettrotoni possono arrivare dai dendriti al soma. Nella regione della sinapsi, conoscendo le leggi sopra discusse, è possibile determinare per quale distanza possono propagarsi i potenziali sinaptici.
Tuttavia, sia i calcoli che le misurazioni dirette forniscono distanze molto piccole su cui un elettrotone può propagarsi. Pertanto, se la depolarizzazione nella regione della membrana non raggiunge il punto CUD, non vi è alcun aumento della permeabilità per il Na+ attraverso i canali voltaggio-dipendenti, si verificano solo cambiamenti passivi nel potenziale di membrana. La costante λ varia da 0,1 a 5 mm. È chiaro che la comunicazione all'interno del sistema nervoso centrale richiede un meccanismo di segnalazione diverso. L'evoluzione lo ha trovato. Questo meccanismo è la propagazione dell'impulso.
Una misura della conduzione di un impulso lungo un assone è la velocità. La velocità di trasmissione dei potenziali d'azione gioca un ruolo significativo nell'organizzazione delle connessioni nel sistema nervoso. Tipicamente, le fibre nervose a conduzione rapida con una velocità di conduzione superiore a 100 m/s servono riflessi rapidi, quelli che richiedono una reazione immediata. Ad esempio, se il posizionamento di un arto non riesce (sei inciampato) per evitare di cadere; nei riflessi protettivi avviati da uno stimolo dannoso, ecc. I riflessi a risposta rapida richiedono un'elevata velocità nelle componenti afferenti e motorie, fino a 120 m/s. Al contrario, alcuni processi non richiedono una risposta così rapida. Ciò vale per i meccanismi che regolano l'attività degli organi interni, dove è sufficiente una velocità di conduzione di circa 1 m/s.
Consideriamo gli eventi legati alla propagazione di AP lungo una fibra nervosa non mielinizzata (a volte erroneamente si dice non mielina, ma questo non ha senso, poiché la mielina non conduce corrente, è una guaina!). L'inizio di un'onda di eccitazione può essere causato dall'attività del recettore (potenziale generatore) o da processi sinaptici. L'AP può anche essere indotto mediante stimolazione elettrica dell'assone. Se lo spostamento locale del potenziale di membrana (depolarizzazione) supera la soglia, raggiunge il punto CUD e attiva i canali del sodio voltaggio-dipendenti, si verifica AP sotto forma di un'onda di depolarizzazione-ripolarizzazione e si propaga lungo la fibra nervosa. Il punto massimo di DP corrisponde alla massima inversione del potenziale di membrana (overshoot). Ciò si traduce in una situazione in cui il PD, propagandosi lungo la fibra, genera davanti ad esso correnti di Hermann, che scaricano la capacità di membrana, avvicinano il potenziale di membrana della sezione successiva della membrana all'ADC, ecc. Dietro di sé, il PD in movimento lascia un'area della membrana che si trova in uno stato di relativa refrattarietà.
Per la propagazione di un potenziale d'azione è necessario che esso generi ogni volta un potenziale elettrotonico nella regione vicina, quella in cui si diffonde, capace di spostare il potenziale di membrana del valore di soglia, cioè dell'ampiezza del potenziale d'azione deve superare più volte la soglia per la sua occorrenza. Viene chiamato il rapporto AP/soglia fattore di garanzia (Aggiorna/Soglia=5..7).
La velocità di movimento dell'elettrotone e del PD lungo le fibre senza polpa è piccola e non supera 1 m/s. Nei calamari, a causa della combinazione di più assoni in uno solo durante l'embriogenesi, che aumenta il diametro totale del conduttore, la velocità dell'impulso in una fibra non mielinizzata può arrivare fino a 25 m/s. Nei mammiferi la velocità aumenta a causa della mielinizzazione degli assoni. L'elevata resistività della mielina porta al fatto che la membrana delle fibre di polpa acquisisce elevata resistenza e bassa capacità. I canali del sodio voltaggio-dipendenti sono concentrati nei nodi di Ranvier, mentre i canali del potassio responsabili della ripolarizzazione sono concentrati nelle regioni di intercettazione. Queste caratteristiche strutturali portano al fatto che salato la conduzione dell'eccitazione ha un'elevata affidabilità e un'elevata velocità, che sono combinate con l'efficienza (gli assoni della polpa per spostare Na + e K + attraverso la membrana costano meno ATPasi sodio-potassio). Una proprietà biofisica distintiva della conduzione saltatoria degli AP è che le correnti sono chiuse attraverso il mezzo intercellulare, che ha una bassa resistenza, e le correnti seguono sia lungo che attraverso la fibra.
La velocità di trasmissione dell'impulso lungo la fibra della polpa dipende dal diametro di quest'ultima per un semplice rapporto
V=K × d, dove d è il diametro e k è una costante.
Per gli anfibi k=2, per i mammiferi k=6.
La lunghezza del tratto di fibra coinvolto nel processo di trasmissione di un PD è pari a L=t × V, dove t è la durata dell'impulso. Questo indicatore è importante da un punto di vista metodologico, poiché la selezione della distanza interpolare degli elettrodi principali (di registrazione) dipende dalla lunghezza della sezione eccitata del nervo.
Nei tronchi nervosi, le singole fibre nervose afferenti e motorie si trovano in uno stato compatto. La conduzione lungo le singole fibre avviene isolatamente da quelle vicine, può diffondersi in due direzioni dal punto di origine, ha una velocità relativamente costante in qualsiasi parte dell'assone (ad eccezione delle terminazioni) e l'eccitazione proviene da diverse fonti di origine nella cellula possono essere soggetti a sommazione algebrica. La gamma di differenze nelle velocità di conduzione nelle fibre è ampia, il che ha consentito di effettuare diverse classificazioni. Le classificazioni più accettate sono Erlanger-Gasser (gruppi AαβγδBC) e, in misura minore, Lloyd (gruppi I, II, III).
L'eccitabilità è la capacità di un tessuto di rispondere alla stimolazione con una reazione specifica per un dato tessuto. L'eccitazione è la transizione del tessuto eccitabile da uno stato di riposo funzionale ad un'attività specifica (per la generazione nervosa e la conduzione degli impulsi nervosi, per il tessuto muscolare - contrazione, per il tessuto ghiandolare - secrezione di secrezioni ghiandolari). In questa sezione esamineremo la struttura e le funzioni del tessuto nervoso e muscolare.
Una cellula nervosa è un anello della catena nervosa. Le singole cellule nervose, o neuroni, non svolgono le loro funzioni come unità isolate, come le cellule del fegato o dei reni. Il lavoro di 50 miliardi di neuroni nel cervello umano è che ricevono segnali da alcune altre cellule nervose e li trasmettono a terze cellule. Le cellule trasmittenti e riceventi sono combinate in circuiti nervosi o reti. Un singolo neurone con un sistema ramificato di processi in uscita (struttura divergente) può inviare il segnale ricevuto in ingresso a mille o anche più altri neuroni. Allo stesso modo, qualsiasi altro neurone può ricevere informazioni di input da altri neuroni utilizzando una, più o moltissime connessioni di input se i percorsi convergenti convergono su di esso.
Vengono chiamate le giunzioni effettive dei neuroni, punti specifici sulla superficie delle cellule in cui avviene il contatto sinapsi. Sostanze chimiche rilasciate nelle sinapsi - neurotrasmettitori- chiudono il circuito tra i neuroni in contatto e sono intermediari chimici per la trasmissione dell'eccitazione nervosa tra le cellule nervose con sinapsi chimiche. Successivamente impareremo che i metodi di trasmissione delle informazioni tra i neuroni possono essere diversi.
1. Un neurone è una cellula.
I neuroni hanno una serie di caratteristiche comuni a tutte le cellule del corpo (Fig. 1).
Riso. 1. Cellula ordinaria (A) e neurone (B).
Come loro, il neurone ha una membrana plasmatica che definisce i confini della singola cellula. Con l'aiuto della membrana e dei meccanismi molecolari in essa contenuti, il neurone interagisce con altre cellule e percepisce i cambiamenti nell'ambiente locale che lo circonda. Con l'aiuto delle proprietà selettive dei canali ionici della membrana e del sistema di trasporto ionico attivo e passivo, la cellula regola la pressione osmotica intracellulare e, quindi, mantiene la sua forma, e garantisce anche che elementi minerali, nutrienti e gas entrino nella cellula per il processo intracellulare bisogni; i potenziali elettrici vengono generati sulla membrana cellulare, alla base della funzione dei neuroni e di altre cellule. La membrana circonda su tutti i lati il contenuto interno della cellula, chiamato citoplasma. Il citoplasma contiene il nucleo e gli organelli citoplasmatici necessari affinché la cellula esista e svolga il suo lavoro: mitocondri, microtubuli, neurofibrille, reticolo endoplasmatico.
I mitocondri forniscono energia alla cellula. Utilizzando zucchero e ossigeno, sintetizzano speciali molecole ad alta energia (ATP), che vengono consumate dalla cellula secondo necessità. I microtubuli, sottili strutture di supporto, aiutano il neurone a mantenere la sua forma e si ritiene siano coinvolti nel trasporto assonale. Con l'aiuto del reticolo endoplasmatico - una rete di tubuli della membrana interna - e delle strutture situate su di esso - i ribosomi - la cellula sintetizza le sostanze necessarie per la sua vita, le distribuisce nella cellula e le secerne oltre i suoi confini. Le neurofibrille svolgono funzioni di supporto, assicurano il movimento direzionale dei terminali degli assoni (rami terminali) e sono uno degli anelli di collegamento nel meccanismo di trasporto degli assoni (vedi sotto).
Il nucleo di un neurone contiene informazioni genetiche codificate nella struttura chimica dei geni. In base a queste informazioni, una cellula completamente formata sintetizza sostanze specifiche che determinano la forma, la chimica e la funzione di questa cellula per tutta la sua vita, poiché la cellula nervosa non è in grado di dividersi.
A differenza delle altre cellule, i neuroni hanno contorni irregolari: hanno processi di diversa lunghezza, spesso numerosi e ramificati. Questi processi sono “fili” viventi con l’aiuto dei quali si formano le reti neurali. Le funzioni di tali fili vengono eseguite anche dalle membrane dei corpi delle cellule nervose.
Le reti nervose degli animali e degli esseri umani presentano una differenza fondamentale rispetto alle reti elettriche tecniche (comprese le reti telefoniche, telegrafiche e Internet): non sono alimentate da generatori centrali di corrente elettrica. Le correnti elettriche si formano sulle membrane molecolari (membrane) delle cellule, a causa della proprietà specifica di queste membrane di generare costantemente potenziali elettrici, e nelle cellule dei tessuti eccitabili (nervi e muscoli) di condurli lungo la loro lunghezza.
Una cellula nervosa ha un processo principale, l'assone, attraverso il quale trasmette le informazioni ad un'altra cellula, e numerosi dendriti, attraverso i quali la cellula riceve le informazioni in arrivo. Sia l'assone che i dendriti possono ramificarsi; Nei dendriti si osserva una ramificazione particolarmente estesa. Sui dendriti e sulla superficie della parte centrale della cellula che circonda il nucleo, chiamata corpo del neurone, si trovano sinapsi di input formate dagli assoni di altri neuroni. Le terminazioni dell'assone o dei suoi rami (terminali) formano tali sinapsi sulla superficie delle membrane di altre cellule. Queste terminazioni contengono organelli chiamati vescicole sinaptiche, in cui si accumulano le molecole del trasmettitore rilasciate dal neurone nella sinapsi della cellula ricevente. Gli assoni mancano di organelli cellulari che sintetizzano le proteine, quindi questo processo, privato della connessione con il corpo cellulare, muore.
I lunghi processi dei neuroni sono chiamati fibre nervose.
2. Conduttore o generatore di corrente elettrica? Funzioni elettrogeniche della membrana cellulare.
Se un microelettrodo viene inserito in una fibra nervosa a riposo e collegato ad un secondo elettrodo inserito sul lato esterno della membrana, il dispositivo di registrazione mostrerà una differenza di potenziale di 60-90 mV, con il lato interno della membrana avente una carica negativa e il lato esterno ha una carica positiva. Questa differenza di potenziale in elettrofisiologia è chiamata potenziale di riposo (RP). Quali forze, separando le cariche su entrambi i lati della membrana, creano la sua polarità elettrica? Una risposta convincente a questa domanda fu data dagli scienziati inglesi A. Hodgkin, A. Huxley e B. Katz (1956-1963), che ricevettero il Premio Nobel per i risultati del loro lavoro
Per comprendere meglio l'essenza della questione, si dovrebbe partire dal fatto che nel corpo degli animali e dell'uomo ogni cellula è circondata da una soluzione di una miscela di sali diversi, principalmente cloruri e fosfati di sodio, potassio, magnesio , ecc., così come i loro composti organici. Gli stessi composti si trovano all'interno della cellula, nel suo citoplasma. E se la cellula fosse passiva a questo fatto, non avrebbe alcuna conseguenza sulla sua vita e attività e non esisterebbero cariche attorno alla membrana, poiché la concentrazione dei sali e, di conseguenza, dei loro ioni su entrambi i lati della sua membrana sarebbe la Stesso. Va notato, tuttavia, che la vita stessa in tal caso sarebbe impossibile.
Ma si è scoperto che la membrana cellulare ha la capacità, utilizzando un sistema di trasporto attivo, di trasportare gli ioni potassio all'interno della cellula e gli ioni sodio verso l'esterno contro un gradiente di concentrazione. Questa capacità è assicurata dall'azione delle pompe potassio-sodio integrate nella membrana, che utilizzano l'energia ATP per il loro lavoro (Fig. 2).

Riso. 2. Schema di funzionamento della pompa ionica.
In un ciclo, la pompa rimuove 3 ioni Na+ dalla cellula e restituisce al suo interno 2 ioni K+, spendendo per questo lavoro l'energia di una molecola di ATP. Naturalmente, la molecola di ATP viene scissa per formare una molecola di ADP e un residuo di fosfato. La cellula crea così quantità squilibrate di sodio e potassio attorno alla sua membrana: all'interno della membrana si crea una concentrazione di ioni potassio 30-50 volte maggiore che all'esterno, e gli ioni sodio nel liquido extracellulare sono 10 volte maggiori che nel citoplasma della cellula. Ciò, tuttavia, di per sé crea una differenza nelle concentrazioni dei cationi, ma non delle cariche.
La seconda parte del processo di polarizzazione (nel tempo avviene contemporaneamente alla prima) fornisce la semipermeabilità o, in altre parole, permeabilità selettiva (selettività) della membrana per vari ioni. A riposo, la membrana lascia passare liberamente gli ioni K +, molto scarsamente gli ioni Na + ed è completamente impermeabile ai grandi anioni di acidi organici e proteine. La disposizione non in equilibrio degli ioni potassio e sodio attorno alla membrana, creata dalle pompe ioniche, provoca un processo passivo: la diffusione di questi cationi lungo un gradiente di concentrazione. All'interno della cellula, gli ioni di potassio si muovono verso la membrana e, superando gli ioni negativi meno mobili, escono verso il suo lato esterno. Di conseguenza, all'interno della membrana viene creata una carica elettrica negativa, mentre all'esterno aumenta la carica positiva creata dagli ioni sodio e la differenza di potenziale calcolata tra loro è di 80-100 mV. La diffusione degli ioni sodio nella cellula è molto più debole (25 volte) di quella del potassio, ma esiste ancora e provoca lo stesso effetto, ma diretto nella cellula, e di conseguenza riduce leggermente il potenziale di potassio calcolato della membrana. Pertanto, il potenziale di membrana a riposo (RMP) risultante è 50-90 mV. (Fig.3)

Riso. 3. Carica elettrica della membrana.
Tali flussi di diffusione di ioni sodio e potassio porterebbero inevitabilmente all'equalizzazione delle concentrazioni di ioni su entrambi i lati della membrana. Ciò, tuttavia, non avviene in una cellula vivente, poiché le pompe ioniche trasportano costantemente ioni contro i gradienti esistenti, mantenendone la concentrazione e i gradienti elettrochimici e, quindi, il potenziale di riposo. Questo è forse uno dei segni funzionali principali e più essenziali della vita. Arrestare il funzionamento delle pompe ioniche nella membrana cellulare significa arrestare tutti i processi vitali, ad es. morte cellulare.
3. La vita delle cellule è elettrizzata. Il ruolo del potenziale di riposo nella vita di una cellula.
Perché una cellula normale ha bisogno di elettricità? È noto che nel corpo degli animali e degli esseri umani, oltre ai nervi e ai muscoli, sono presenti cellule della pelle, organi interni e tessuti circostanti. Si è scoperto che tutti hanno potenziali di membrana rispettivamente da 15 a 50 mV. Pertanto, le cellule nervose non hanno il monopolio per quanto riguarda la presenza di potenziali elettrici e, quindi, l'uso dell'elettricità nelle loro attività vitali. Perché le cellule hanno bisogno di elettricità? Proviamo a immaginare come sarebbe se chiedessimo, ad esempio, a una casalinga, perché ha bisogno dell'elettricità? Oppure perché qualsiasi moderna impresa di produzione industriale, situata, ad esempio, al secondo piano di un edificio, ha bisogno di elettricità?
Oltre all'acqua e ad alcuni elettroliti, che passano liberamente attraverso la membrana cellulare, la cellula ha bisogno di sostanze nutritive, ad esempio zuccheri, amminoacidi e alcuni elementi minerali che non possono penetrare nella cellula secondo le leggi della diffusione. In realtà, nel processo di evoluzione, la cellula potrebbe acquisire una membrana che consentirebbe il libero passaggio di tutti gli ioni e le molecole di sostanze organiche e tutti i problemi relativi alla fornitura di nutrienti alla cellula verrebbero risolti secondo le leggi della diffusione. Perché ciò non è realmente accaduto? In primo luogo, secondo le leggi della diffusione, gli ioni e le molecole delle sostanze si sposterebbero nella cellula solo se fossero più concentrati all'esterno; in secondo luogo, l'attività vitale della cellula non tollera le interruzioni che potrebbero verificarsi nell'approvvigionamento della sostanza necessaria; di tanto in tanto a causa della sua diminuzione nell'ambiente cellulare circostante, in terzo luogo, l'allineamento su entrambi i lati della membrana cellulare di prodotti metabolici dannosi per la cellula non solo avrebbe di per sé un effetto dannoso sulla cellula, ma avrebbe anche ridurre la concentrazione delle sostanze necessarie per la cellula nell'ambiente esterno, provocandone così il deflusso inverso dalla cellula. Gli interessi vitali della cellula si rivelarono incompatibili con tale elemento “di mercato”, e l’evoluzione della cellula rese la sua membrana selettivamente permeabile ad alcuni ioni e la dotò di mezzi di trasporto attivi per “importare” altre sostanze nella cellula, nonché per “esportare” i prodotti metabolici di cui non ha bisogno dalla cellula. La cellula non doveva scegliere il tipo di energia per garantire il funzionamento dei suoi mezzi di trasporto: conosceva l’elettricità milioni di anni prima dello scienziato greco Talete, che indicò la capacità dell’ambra strofinata con la seta di attrarre oggetti leggeri,
Come veicoli di trasporto, le molecole proteiche trasportatrici sono integrate nella membrana cellulare e trasportano i nutrienti nella cellula come segue (Fig. 4).

Riso. 4. Schema di trasporto di membrana.
All'esterno della membrana, la molecola della proteina trasportatrice attacca a sé una molecola nutritiva e uno ione sodio, acquisendo così una carica positiva. Il campo elettrico della membrana, ruotando la molecola portatrice attorno al suo ipotetico asse, attira il suo polo carico positivamente verso l'interno della membrana. Qui lo ione sodio perde la sua carica e lascia la cellula trasportatrice, il che porta al distacco da essa di una molecola neutra della sostanza nutritiva trasportata. Il nutriente rimane all'interno della cellula e il sodio viene pompato fuori, ripristinando una carica positiva all'esterno della membrana.
Esistono diverse proteine trasportatrici per trasportare diversi zuccheri e amminoacidi nella cellula, ma tutte utilizzano gli ioni sodio e il potenziale di membrana a riposo nelle cellule animali. I batteri utilizzano ioni idrogeno invece del sodio
Il trasporto elettrico viene utilizzato per rimuovere determinate sostanze dalla cellula. Il calcio in eccesso nella cellula, se la pompa del calcio non riesce a far fronte alla sua rimozione, è pericoloso. Il sistema di emergenza della gabbia si attiva. In questo caso, una speciale proteina trasportatrice lega uno ione calcio alla sua parte intracellulare e tre ioni sodio alla sua parte esterna. Il campo elettrico della membrana ruota la molecola trasportatrice di 180° e il calcio viene rilasciato. A differenza delle pompe potassio-sodio, che utilizzano l'energia dell'ATP per il loro lavoro e allo stesso tempo creano un campo elettrico (potenziali elettrici) sulla membrana, le suddette proteine trasportatrici funzionano come motori elettrici che utilizzano l'energia elettrica del potenziale di membrana per il loro lavoro.
I meccanismi di trasferimento attivo dei nutrienti nella cellula sono necessariamente associati alla sua vita e, senza dubbio, avrebbero dovuto sorgere insieme all'emergere della cellula. La trasmissione dei segnali da cellula a cellula è una successiva esigenza dell'evoluzione, quindi è naturale supporre che quando si è presentata tale necessità, le cellule nervose, avendo in qualche modo modificato la loro membrana, hanno utilizzato il potenziale della membrana per una nuova funzione: la trasmissione del segnale. In cosa consisteva questa modifica?
4. Cambiamenti nel potenziale di membrana. Potenziale d'azione.
Nel 1939, i dipendenti della stazione marittima di Plymouth (Inghilterra) A. Hodgkin e H. Huxley misurarono per la prima volta il PP e l'AP di una fibra nervosa, utilizzando un assone di calamaro gigante (1 mm di diametro) e il più avanzato strumento sperimentale tecnologia in quel momento. Si è scoperto che a riposo sulla membrana la differenza di potenziale è di circa 80 mV, e quando eccitata la membrana non si scarica, come ipotizzava Bernstein, ma si ricarica: le sue cariche negative sono all'esterno e le cariche positive sono all'interno. In questo caso sulla membrana appare una differenza di potenziale di circa 40 mV. Ciò significa che durante l'eccitazione il potenziale non cambia del valore del PP, cioè non scende a zero, ma dalla somma del PP e del nuovo potenziale emergente con segno opposto (Fig. 8). Questo eccesso di PD rispetto a PP è stato chiamato overshoot (inglese oversoot – eccesso).
Per spiegare questo potenziale “aggiuntivo”, Hodgkin e Huxley avanzarono un'ipotesi, la cui essenza era che quando eccitata, la membrana non solo perde completamente la sua permeabilità selettiva, ma la cambia: da permeabile principalmente agli ioni K +, la membrana diventa permeabile principalmente agli ioni Na + . E poiché questi ultimi sono più numerosi all'esterno della membrana, si precipitano all'interno e ricaricano la membrana.
Il primo test dell’ipotesi fu intrapreso da A. Hodgkin e B. Katz dopo una lunga pausa dovuta alla guerra, nel 1949. Gli studi hanno dimostrato che l'ampiezza AP dipende dai cambiamenti nella concentrazione degli ioni Na+ nello stesso modo in cui il valore PP dipende dalla concentrazione degli ioni K+, cioè obbedisce alla formula di Nernst.
Successivamente, è stato necessario scoprire come cambia effettivamente la permeabilità della membrana al potassio e al sodio durante l'eccitazione e da quali condizioni dipendono questi cambiamenti. Hodgkin e Huxley hanno proposto che la permeabilità della membrana sia determinata da due condizioni: il potenziale di membrana e il tempo trascorso dalla variazione di potenziale. Il risultato di una lunga e laboriosa verifica di questa ipotesi è stata la conferma sperimentale che il meccanismo di tutti i fenomeni associati all'eccitazione si basa effettivamente sulla proprietà della membrana: la sua permeabilità selettiva variabile per gli ioni potassio e sodio. È stata creata una teoria per la comparsa della PD in diverse condizioni e stati iniziali della membrana, ad es. “teoria moderna delle membrane”, chiamata anche modello X-X. Il modello riproduceva regolarmente fenomeni come la refrattarietà, la soglia di eccitazione, l’iperpolarizzazione delle fibre dopo un impulso, ecc.
Tuttavia, c'erano ancora domande senza risposta: qual è il meccanismo che modifica la permeabilità della membrana, come è strutturata questa partizione più sottile? La penetrazione nei segreti della struttura della membrana e dei suoi meccanismi funzionali iniziò più tardi, quando apparve una nuova scienza: la biologia molecolare.
Struttura della membrana. La membrana citoplasmatica è un film molecolare passivo costituito da due strati di molecole lipidiche; contiene molecole proteiche che svolgono funzioni attive nella membrana come canali e pompe ioniche, molecole trasportatrici, recettori, ecc. Lo strato bimolecolare di lipidi isola la cellula dall'ambiente ed è anche un isolante elettrico. La connessione della cellula con l'ambiente e tutte le funzioni attive della sua membrana vengono eseguite utilizzando molecole proteiche integrate nella membrana (Fig. 5)

Riso. 5. Membrana plasmatica cellulare
Alcune molecole proteiche forniscono la capacità di sensibilità locale della membrana agli effetti degli stimoli e la risposta adeguata a questi effetti. La modifica evolutiva della membrana delle cellule nervose consisteva nella creazione sulla membrana di meccanismi per cambiamenti locali nei potenziali di membrana con la successiva propagazione ondulatoria di questo cambiamento lungo la membrana. Quest'ultima capacità è posseduta anche dalle membrane delle cellule di altri tessuti.
Canali ionici. Le proprietà elettriche attive della membrana cellulare sono determinate dalle molecole proteiche in essa integrate, che formano “pori” in una membrana lipidica omogenea e impenetrabile per molecole e ioni, attraverso i quali queste particelle possono passare in determinate condizioni. I pori attraverso i quali possono passare gli ioni potassio e sodio sono chiamati rispettivamente canali ionici potassio e sodio.
I canali ionici sono formati da una classe speciale di molecole proteiche che sono in grado di distinguere i “loro” ioni e di aprire o chiudere loro il percorso attraverso la membrana sotto l'influenza del potenziale di membrana appropriato. Il biofisico inglese B. Hill ha scoperto che il diametro del canale del potassio è di circa 0,3 nm e il canale del sodio ha un lume leggermente più grande per il passaggio degli ioni. A differenza del canale K+ del sodio, è privo di un meccanismo di inattivazione. Va notato che il passaggio dell'uno o dell'altro al canale è determinato non dal diametro di quest'ultimo, ma dalle sue proprietà funzionali e dai potenziali elettrici della membrana.
Il canale ionico può essere pensato come un tubo passante integrato nella membrana (Fig. 6)

Riso. 6. Schema di funzionamento del canale ionico.
In prossimità dell'estremità esterna, il tubo ha un “otturatore” o “porta”, la cui posizione è controllata dal potenziale di membrana. L'“otturatore” si carica e quindi, quando cambia il potenziale (durante la depolarizzazione), può aprire la membrana ingresso nel canale per lo ione corrispondente. Si ritiene che le porte del canale siano un gruppo carico di atomi di una molecola proteica che può muoversi in un campo elettrico, aprendo così la strada agli ioni potassio o sodio. Lo spostamento di un gruppo così carico deve essere accompagnato da una piccola corrente elettrica a breve termine; Questa corrente nei canali del sodio è stata registrata sperimentalmente ed è stata chiamata “corrente di gate”). Studiando i canali del sodio, è stato dimostrato che i meccanismi di gate e di inattivazione si trovano a diverse estremità del canale. Dopo aver rimosso l'estremità interna del canale utilizzando l'enzima pronasi, il canale ha continuato ad aprire la porta del sodio sotto depolarizzazione, ma non è stato inattivato.
È stata determinata la densità dei canali del sodio nella membrana. Ce n'erano circa 50 per micrometro quadrato di membrana (quest'area della membrana può ospitare diversi milioni di molecole lipidiche).
Oltre ai canali del potassio e del sodio, le membrane cellulari hanno canali per il calcio, il cloro e altri ioni. Un cambiamento graduale nella permeabilità della membrana, ad esempio per gli ioni potassio, è spiegato dal fatto che i canali corrispondenti, oltre agli stati "aperto" e "chiuso", hanno alcuni altri stati, ad es. che i canali sono caratterizzati da un funzionamento probabilistico, confermato sperimentalmente. Inizialmente si presumeva che il canale ionico fosse un meccanismo biologico stabile che è integrato nella membrana e funziona per un periodo piuttosto lungo, ma in seguito si è scoperto che le proteine del canale funzionano solo per circa un giorno, quindi vengono smantellate e sostituiti da altri sintetizzati dai ribosomi della cellula su comando dell'RNA messaggero. Si è scoperto che le proteine canale non vengono inserite in modo casuale nella membrana e che i diversi canali ionici vengono trasportati dalla cellula esattamente nel posto giusto.
Oltre ai canali la cui apertura e chiusura è controllata dal potenziale elettrico della membrana, esistono canali ionici controllati da sostanze chimiche (non solo trasmettitori nelle sinapsi) che agiscono sulla membrana sia all'esterno che all'interno della cellula. Ad esempio, un aumento della concentrazione di adenosina monofosfato ciclico (cAMP) in alcuni neuroni porta all'apertura dei canali e alla depolarizzazione della cellula, e un aumento di questa concentrazione è stimolato dall'adrenalina. Una cellula non solo può controllare il funzionamento dei canali ionici, ma può modificarli e cambiarne le proprietà attraverso reazioni biochimiche controllate. Tali processi si verificano, ad esempio, durante l'apprendimento.
Cambiamenti nel potenziale di membrana . La membrana cellulare ha la proprietà di rispondere all'irritazione modificando il potenziale di riposo, un processo in rapido aumento della sua depolarizzazione locale. L'entità di questa depolarizzazione dipende dall'intensità dello stimolo e nelle cellule nervose e muscolari può provocare una completa scomparsa a breve termine del potenziale di membrana nell'area di stimolazione, seguita da un'inversione di polarità e quindi da una ripolarizzazione, cioè ripristino dello stato originario. Questo fenomeno è chiamato potenziale d'azione. Il livello del potenziale di membrana al quale un ulteriore aumento dell'intensità dello stimolo e un'ulteriore depolarizzazione danno origine a un potenziale d'azione è chiamato soglia, e lo stimolo corrispondente è chiamato soglia.
Livelli di depolarizzazione della membrana. Se la forza dello stimolo che agisce sulla membrana non supera lo 0,5 del valore soglia dello stimolo, la depolarizzazione della membrana verrà notata solo durante l'azione dello stimolo. Questo fenomeno è chiamato potenziale elettrotonico (Fig. 7)

Riso. 7. Potenziale elettrotonico.
La permeabilità ionica della membrana rimane praticamente invariata. Quando l'intensità dello stimolo aumenta fino a 0,9 del valore soglia, il processo di depolarizzazione segue una curva a forma di S, continua ad aumentare per qualche tempo dopo la cessazione dello stimolo, ma poi si arresta lentamente. Questa reazione della membrana cellulare è chiamata risposta locale. Una risposta locale non è in grado di propagarsi. Tuttavia, quando vengono applicate più stimolazioni sottosoglia ad un intervallo più breve della durata di una risposta locale individuale, queste ultime si sommano e aumenta la depolarizzazione della membrana, che può portare ad un potenziale d'azione nelle cellule nervose e muscolari. Le cellule epiteliali non possono generare un potenziale d'azione. La loro reazione a uno stimolo di qualsiasi intensità è limitata a una risposta locale.
Quando uno stimolo di forza soglia o soprasoglia viene applicato alla membrana, il potenziale di membrana diminuisce di 20-40 mV rispetto al potenziale di riposo, cioè ad un certo valore critico. Se la forza dello stimolo non diminuisce, i pori (canali) nella membrana per gli ioni sodio si aprono bruscamente e la sua permeabilità per questi cationi aumenta decine, anche centinaia di volte. Lungo la concentrazione e i gradienti elettrochimici, gli ioni sodio si riversano nella cellula come una valanga, portando con sé le cariche positive. Sul lato esterno della membrana, nella zona eccitata, invece delle cariche positive, si formano cariche negative dovute a grandi anioni di acidi organici e proteine. Il lato interno della membrana percepisce questo flusso a valanga di cationi Na come un'iniezione di corrente elettrica con segno positivo.
L'accumulo di ioni Na+ sul lato interno della membrana porta prima alla neutralizzazione della sua carica negativa, cioè alla sua depolarizzazione, e quindi all'instaurazione di una carica positiva in questo luogo: la depolarizzazione è sostituita dall'inversione di carica (Fig. 8).

Riso. 8. Potenziale d'azione.
Il potenziale di membrana nel sito dello stimolo diventa positivo. Questa fase positiva del potenziale di membrana è chiamata superamento o picco e l'altezza sulla curva del potenziale è 40-50 mV. Graficamente, questo è il ramo ascendente della curva del potenziale d'azione. In questo momento si verifica l'inattivazione dei canali del sodio, la permeabilità della membrana agli ioni sodio cessa, lasciando il posto ad un aumento della conduttività del potassio. L'aumento del flusso di ioni potassio verso l'esterno ripristina la polarità originale, provocando la ripolarizzazione della membrana, e le pompe del sodio che si accendono successivamente, pompando gli ioni sodio fuori dalla cellula, completano il processo di ripolarizzazione.

Riso. 9. Cambiamento nell'eccitabilità della membrana delle fibre nervose
durante un potenziale d'azione.
Graficamente questi processi sono caratterizzati dal ramo discendente della curva del potenziale d'azione. Il flusso di ioni Na+ verso il lato esterno della membrana porta ad un leggero aumento del potenziale di membrana rispetto a quello iniziale. Questo è il cosiddetto "traccia iperpolarizzazione" della membrana. Durante questo periodo, chiamato periodo refrattario, la membrana è insensibile alla stimolazione ripetuta. Questo fatto ha due conseguenze. In primo luogo, l'insensibilità di questa sezione della membrana all'irritazione ripetuta non consente il ritorno dell'impulso dalla sezione vicina e, in secondo luogo, le correnti elettriche ad alta frequenza non causano eccitazione (potenziali d'azione) nel nervo, ma riscaldano solo la parte corrispondente aree dei tessuti (e anche il nervo stesso).
L'ampiezza della curva del potenziale d'azione è determinata come la somma dei valori del potenziale di membrana a riposo e del suo eccesso - superamento dopo l'inversione di carica. Ad esempio, con un PP di -80 mV e un superamento di +50 mV, l'ampiezza del PD sarà di 130 mV e i segni dei suoi componenti non verranno presi in considerazione. La durata dell'AP sulla membrana delle fibre nervose è 1-3 ms, i muscoli - fino a 8 ms, il muscolo cardiaco - 300 ms.
Inattivazione del sistema Na+ . Nell'assone del calamaro, con depolarizzazione costante, la permeabilità della membrana agli ioni Na + inizia a diminuire dopo 0,5 ms nei neuroni dei vertebrati questo periodo è 5 volte inferiore; Questa rapida diminuzione della permeabilità della membrana agli ioni sodio è chiamata inattivazione. La massima permeabilità della membrana agli ioni sodio si osserva ad un livello di depolarizzazione circa 30-40 mV più negativo del potenziale di riposo. A potenziali iniziali 20-30 mV più positivi del potenziale di riposo, il sistema Na+ è completamente inattivato e non può essere attivato da alcuna depolarizzazione.
L'inattivazione voltaggio-dipendente del sistema Na influisce in modo critico sull'eccitabilità cellulare in varie condizioni. Pertanto, se il PP delle cellule di mammifero diventa più positivo di -50 mV (ad esempio, a causa della mancanza di ossigeno o sotto l'influenza di miorilassanti), la conduzione del sodio viene completamente inattivata e la cellula diventa ineccitabile. Quando la concentrazione di Ca 2+ aumenta, la cellula diventa meno eccitabile e quando diminuisce aumenta l'eccitabilità della cellula. Questo aumento di eccitabilità è alla base delle sindromi tetaniche e di altre condizioni associate a una carenza di calcio ionizzato nel sangue; in questo caso si verificano contrazioni muscolari involontarie e crampi.
5. Movimento di eccitazione lungo la fibra nervosa.
Secondo i concetti moderni, la diffusione dell'eccitazione lungo le fibre nervose e muscolari è spiegata dalla generazione di potenziali d'azione sulle loro membrane e dall'impatto delle correnti elettriche locali risultanti sulle aree vicine della membrana. Quando un potenziale d'azione si verifica in qualsiasi parte della membrana durante la fase ascendente della curva AP fino al suo picco (superamento), questa parte della membrana diventa un generatore biologico di corrente elettrica e al suo interno scorre una corrente di segno positivo dall'esterno della membrana nella cellula. La fonte della corrente elettrica è il rapido flusso di ioni sodio attraverso la membrana.
Sul lato interno della membrana, una corrente elettrica di segno positivo, neutralizzandone le cariche negative, scorre verso le zone non eccitate, vincendo la resistenza del citoplasma, e contemporaneamente scorre verso il suo lato esterno, dove già scorre verso la zona eccitata, utilizzando il fluido extracellulare come un filo (Fig. 9, A).
Queste correnti utilizzano il citoplasma e la membrana della cellula come un filo elettrico passivo scarsamente isolato posto in un ambiente elettricamente conduttivo e sono chiamate elettrotonico correnti. Finché l'intensità di queste correnti è inferiore alla soglia, non generano potenziale d'azione e vengono anche chiamate correnti locali. Ma se la loro forza, fino ad una certa distanza dal luogo di generazione, ad esempio dal luogo di origine del precedente AP, è parecchie volte superiore al valore di soglia, allora tali correnti stimolano la depolarizzazione della sezione vicina di membrana con la conseguente comparsa qui di un potenziale d'azione. In futuro, tutto ciò si ripeterà nel prossimo sito vicino; l'eccitazione si muove in onde lungo la membrana. Pertanto, quando si trasmette un segnale lungo una fibra nervosa, si verificano due processi interagenti. Il primo processo è l'eccitazione della membrana, cioè la generazione di PD è dovuta a fattori specifici, ad es. proprietà elettrogeniche attive della membrana delle cellule eccitabili. La seconda - la trasmissione del segnale lungo la fibra dalla sezione eccitata a quella non eccitata - è puramente elettrica ed è dovuta alle sue proprietà elettriche passive, quando la fibra nervosa si comporta come un conduttore passivo di potenziali elettrici. La conduzione dell'eccitazione lungo una fibra nervosa utilizzando il primo processo è chiamata pulsata, il secondo - non pulsata o elettrotonica. Il metodo di trasmissione senza polso avviene in determinate reti nervose, dove è richiesta la sua trasmissione su determinate distanze abbastanza brevi (vedi sotto). Durante la conduzione dell'impulso, una fibra nervosa funziona come una catena di dispositivi relè collegati da un conduttore attraverso il quale i potenziali elettrici viaggiano da relè a relè. Tuttavia, poiché una fibra nervosa può trasmettere un segnale in modo puramente elettrico, allora, in primo luogo, perché ha bisogno di ripetitori intermedi e, in secondo luogo, se sono ancora necessari, quanto dovrebbero essere lunghe le distanze tra i punti di trasmissione della fibra nervosa?
Costante di lunghezza delle fibre nervose . A differenza di un cavo con un nucleo metallico situato nell'aria, in cui la tensione viene spesa solo per superare la resistenza del nucleo stesso, in una fibra nervosa la tensione viene spesa sulla resistenza del "nucleo" del cavo - l'assoplasma, il suo isolamento: la membrana e la dispersione di corrente attraverso i pori della membrana nel fluido extracellulare. Pertanto, la potenza del segnale iniziale diminuisce gradualmente e il segnale svanisce. La lunghezza della sezione della fibra nervosa in cui la forza attuale derivante dal sito di eccitazione diminuisce di e volte è chiamata costante di lunghezza ed è indicata con la lettera greca l-lambda. La lettera e denota la base dei logaritmi naturali, che è circa 2,718. Nelle cellule nervose, la costante di lunghezza della fibra nervosa varia da 0,1 a 5 mm. Ad esempio, a una distanza di 4 lambda, l'ampiezza del potenziale elettrotonico è solo il 2% del potenziale originale. Pertanto, in una fibra nervosa a una distanza di 1 m, non solo il potenziale elettrotonico, ma anche il segnale causato da AP si estinguerà completamente. Pertanto, affinché il segnale possa propagarsi alla sua forza originale, deve essere amplificato. Ma la lunghezza della fibra nervosa non raggiunge sempre 1 m. Nei piccoli insetti, la lunghezza costante della fibra nervosa è di 250 micron, che nella Drosophila rappresenta il 20% del suo corpo. E a questa distanza il segnale originale si attenuerà solo 2.718 volte. Questa quantità di attenuazione non interferisce con la trasmissione del segnale. Ma una tale lunghezza è caratteristica di molti neuroni nel cervello dei vertebrati. Negli animali superiori ci sono cellule nervose aperte che non generano AP lungo la loro lunghezza, ma sono conduttori passivi di potenziali elettrotonici (ad esempio i neuroni della retina).
Tempo costante . Quindi, per caratterizzare la distanza sulla quale un segnale può essere trasmesso lungo le fibre nervose non pulsative, ad es. metodo elettrotonico, ogni volta che è necessario prendere un'unità di lunghezza: la costante di lunghezza o costante di attenuazione ( l) fibra nervosa.
Tuttavia la costante di smorzamento è utile solo in caso di corrente continua. Ma nessuna informazione può essere trasmessa utilizzando la corrente continua; Ciò richiede corrente alternata. Quando si determina la velocità di propagazione di un segnale lungo una fibra nervosa che coinvolge corrente alternata, è necessario tenere conto del fatto che la sua membrana ha una capacità elettrica (Fig. 10), che deve essere scaricata prima che raggiunga l'ampiezza di picco della corrente alternata il suo massimo.
Per questo è necessario dedicare una certa quantità di tempo. Durante questo periodo, la corrente non solo scarica la capacità di questa sezione della membrana, ma fuoriesce anche (nello spazio intercellulare). Di conseguenza, con una corrente alternata che cambia rapidamente, l'efficienza di scarica della membrana sarà inferiore, quindi il segnale si attenua più velocemente.
Il tasso di attenuazione del segnale in questo caso è determinato utilizzando il cosiddetto. costante di tempo (t), pari al prodotto della resistenza (Rm) della membrana e della sua capacità specifica (Cm), cioè
t=RmCm La costante di tempo del potenziale elettrotonico è determinata dal tempo durante il quale il potenziale raggiunge il livello 1/e, cioè 37% della sua ampiezza finale; t=10 ms significa che per una data fibra nervosa l'attenuazione di un segnale della durata di 10 ms è proporzionale alla sua costante di lunghezza (l). Un segnale del genere in questa fibra verrà trasmesso praticamente come una corrente continua, ma per una fibra nervosa con t = 0,5 s (neurone del mollusco) questo segnale sarà già “veloce”, cioè svanirà su una distanza più breve. Nei vertebrati, in diverse cellule la costante di tempo (t) varia da 5 a 50 ms.
Pertanto, per la trasmissione di un segnale sufficientemente lento rispetto alla costante di tempo (t) e su una distanza paragonabile alla costante di attenuazione (l), il metodo di trasmissione del segnale elettrotonico (senza impulsi) è molto adatto. Se la distanza su cui deve essere trasmesso il segnale è significativamente maggiore della costante di attenuazione, è necessaria la sua amplificazione, ad es. trasmissione con generazione sequenziale di potenziali d'azione – trasmissione di impulsi di segnali. Evolution seleziona tra questi metodi quello che meglio garantisce la trasmissione di un segnale di forza sufficiente all'estremità ricevente della fibra nervosa, oppure utilizza entrambi i metodi contemporaneamente, garantendo così l'affidabilità della segnalazione.
Tuttavia, per garantire un funzionamento efficace della fibra nervosa, è necessaria non solo l'affidabilità, ma anche la trasmissione tempestiva del segnale, che richiede una velocità di trasmissione sufficientemente elevata.
Velocità di trasmissione elettrotonica. Questo concetto fu introdotto nel 1946 da A. Hodgkin e V. Rushton]. Prima di tutto era necessario scoprire cosa si dovrebbe intendere per velocità di un elettrotone? Se intendiamo il tempo durante il quale un segnale arbitrariamente piccolo appare in una fibra nervosa ad una certa distanza dal luogo di stimolazione, allora questa sarà la velocità della luce. Ma un segnale di così bassa intensità non può essere ricevuto, poiché nel punto di ricezione deve avere un'intensità di soglia, e quindi un segnale di tale intensità e la velocità indicata dell'elettrotone sono inutili.
Caratteristiche di una fibra nervosa come cavo, la cui membrana è una catena di capacità elettriche (Fig. 9, a e 10,

Riso. 10. Diffusione dell'eccitazione lungo la fibra nervosa.
mostra che la propagazione dei potenziali di membrana lungo una fibra nervosa avviene mediante scarica sequenziale di queste capacità, per la quale è richiesta una corrente di intensità sufficiente in ciascuna sezione della fibra; al punto successivo, una corrente di tale intensità può essere raggiunta solo dopo che la capacità precedente è stata scaricata, e man mano che si scaricano successivamente capacità sempre più distanti, la distribuzione del potenziale si avvicina ad un certo valore stazionario (Fig. 11).

Riso. 11. La velocità di propagazione dell'eccitazione lungo la fibra nervosa.
Nel grafico, questo valore è espresso da una curva esponenziale, che mostra che la velocità con cui un segnale raggiunge un certo punto della fibra che può essere ricevuto dipende dalla velocità con cui lo schema di distribuzione del potenziale lungo la fibra nervosa si avvicina ad un valore stazionario . Questa velocità dipende dalla costante di tempo ( T) e costanti di smorzamento ( l). Più T, minore è la frazione di corrente che fluisce attraverso la membrana nel fluido intercellulare che circonda la fibra per unità di lunghezza della membrana e, quindi, più velocemente la corrente rimanente scaricherà la capacità successiva, e maggiore è la capacità (e quindi l), tanto più lenta sarà la sua scarica e, di conseguenza, l'avvicinamento della curva potenziale in questo tratto ad un valore stazionario.
La curva esponenziale in Fig. 2-11 mostra che è impossibile determinare la velocità dal momento in cui ad un certo punto viene raggiunto un valore potenziale stazionario, perché il tempo per realizzarlo tende all'infinito. Pertanto, Hodgkin e Rushton scelsero una definizione convenzionale della velocità dell'elettrotone. Per fare ciò, hanno utilizzato un esponente dimezzato dei valori stazionari del potenziale di membrana (in Fig. 2-11 è indicato da croci). Lo si può vedere dal punto di partenza UN il potenziale diventa pari a 0,5 del valore stazionario prima che in altri punti della fibra, e successivamente questo tempo aumenta uniformemente di 0,5 T a distanza di l Pertanto, la velocità di trasmissione del segnale elettrotonico è la velocità con cui la fibra nervosa si propaga per raggiungere la metà del valore di stato stazionario del potenziale di membrana. Si esprime con la formula: V= 2 l/T, cioè. pari al doppio della costante di lunghezza della membrana divisa per la sua costante di tempo.
Se accettiamo l=2,5 mm e T=50 ms, allora la velocità dell'elettrotone sarà di 100 mm al secondo.
Per la propagazione elettrotonica di un segnale in una fibra nervosa reale è necessario tenere presente un dettaglio importante: l'ampiezza del potenziale di membrana iniziale deve essere inferiore alla soglia. Il valore limite del potenziale di membrana porta immediatamente la membrana in uno stato di eccitazione, cioè ne provoca la depolarizzazione con la successiva generazione di un potenziale d'azione, che è ciò che osserviamo nelle normali fibre nervose].
Conduzione degli impulsi di eccitazione . La conduzione di un impulso nervoso lungo una fibra nervosa avviene come risultato di due processi alternati: attivo - eccitazione della membrana e passivo - trasmissione del potenziale elettrotonico lungo una sezione non eccitata della fibra. Pertanto, la velocità di conduzione dell'impulso lungo la fibra è un valore composito e consiste nella velocità dell'elettrotone e nel tempo impiegato per generare potenziali d'azione. Maggiore è la velocità dell'elettrotone, più velocemente e lontano il segnale si propagherà lungo la fibra ancora non eccitata, mantenendo un valore soprasoglia, cioè un valore capace di provocare la generazione di un potenziale d'azione. Questo a sua volta dipende dalle caratteristiche attive della fibra: il fattore di sicurezza e la velocità di generazione degli impulsi.
Il fattore di sicurezza è il rapporto tra l'ampiezza del potenziale d'azione e la soglia di eccitazione della fibra. È chiaro che maggiore è l'ampiezza dell'AP e più bassa è la soglia di eccitazione della membrana, maggiore è la distanza in cui l'AP è in grado di attivare la porzione non eccitata della membrana. E, infine, quanto più velocemente procedono i processi di generazione dei PD, tanto più breve è il ritardo per la ritrasmissione e, quindi, tanto maggiore è la velocità di esecuzione.
Quali opportunità ha avuto l'evoluzione per aumentare la velocità di eccitazione lungo la fibra nervosa?
L'ampiezza AP non può essere aumentata in modo significativo per due importanti ragioni. In primo luogo, per questo sarebbe necessario aumentare di centinaia di volte la differenza di concentrazione degli ioni potassio all’interno e all’esterno della fibra e, di conseguenza, aumentare i costi energetici delle pompe ioniche, che sarebbero sproporzionati rispetto ai risultati ottenuti, e, in secondo luogo , trascurano le capacità della membrana, che in generale non esiste essere vivente in natura. Il punto qui è questo. L'ampiezza PD è di circa 0,1 V. Considerando lo spessore della membrana (10 nm), l'intensità di campo su di essa è di 109 V. Se l'ampiezza PD viene aumentata più volte, la membrana verrà perforata. È anche impossibile ridurre significativamente la soglia di eccitazione, perché deve essere superiore al livello delle fluttuazioni naturali del potenziale di membrana a riposo, il cosiddetto. “rumore” della membrana associato al movimento irregolare degli ioni K+ che fluiscono attraverso la membrana.
Pertanto, l'evoluzione ha due modi per aumentare la velocità di propagazione dell'eccitazione nelle fibre nervose: modificandone il diametro o aumentando la distanza tra i punti di generazione AP, ad es. stazioni di ritrasmissione delle fibre nervose.
Abbiamo visto sopra che la velocità di propagazione elettrotonica dei potenziali è proporzionale alla radice quadrata del diametro, cioè che quando il diametro della fibra aumenta di quattro volte, la velocità di propagazione del segnale attraverso di essa raddoppierà: con un diametro della fibra di 200 micron, la velocità è di 20 m/s, con un aumento del diametro della fibra a 2 mm, questa velocità aumenterà aumentare a 60 m/s. Quest'ultima, però, in un organismo complesso porterebbe ad un ingombro dei tessuti con fibre nervose di grande diametro, il che non solo sarebbe antieconomico, ma irrazionale dal punto di vista della forma e della funzione di questi tessuti. Pertanto, l'evoluzione ha scelto il secondo metodo: il metodo di conduzione saltarica dell'eccitazione tra stazioni di trasmissione delle fibre nervose isolate l'una dall'altra. Nelle fibre mielinizzate sottili con un diametro di 20 micron, la velocità di trasmissione dell'impulso raggiunge i 120 m/s. Come si ottiene un tale aumento della velocità di conduzione?
La fibra mielinizzata differisce dalle fibre ordinarie in quanto i suoi tratti piuttosto lunghi sono ricoperti da un buon isolante (mielina). Queste aree mielinizzate si alternano regolarmente ad aree corte (2 µm) non isolate, le cosiddette. nodi di Ranvier, in cui la membrana fibrosa è priva di isolamento. Le sezioni d'intercettazione sono circa 100 volte il diametro di una data fibra; L'isolamento è formato da cellule ricche di mielina che, come un nastro isolante, avvolgono più volte la fibra nervosa, rendendo impossibile il flusso degli ioni attraverso la membrana in queste aree. Nelle intercettazioni la membrana è specializzata nella generazione di potenziali d'azione ed è una sorgente di corrente; la densità dei canali Na + qui è 100 volte maggiore rispetto alla fibra senza polpa (Fig. 12).

Riso. 12. Intercettazioni di Ranvier.
La corrente generata nell'intercettazione raggiunge l'intercettazione successiva senza grandi perdite e, uscendo lì, eccita la generazione di PD sulla sua membrana, che è la fonte di corrente per la successiva sezione interintercettore. Pertanto, quando il potenziale di membrana si sposta, la corrente non scorre attraverso la membrana delle aree interintercettate e l'impulso in queste aree si propaga elettrotonicamente, con un consumo di tempo quasi nullo (0,05-0,07 ms) e quasi senza attenuazione (senza decremento). . Il ritardo di conduzione si verifica solo nelle intercettazioni in cui il potenziale elettrotonico deve raggiungere un'intensità di soglia e causare la generazione di un potenziale d'azione. Questa disposizione della fibra nervosa non solo garantisce un'elevata velocità di eccitazione, ma è anche molto economica: le pompe ioniche funzionano solo nelle intercettazioni
La natura ha scelto la lunghezza del tratto intercetta pari a 100 volte il diametro di questa fibra. Pur non essendo teoricamente il più ottimale, garantisce la conduzione di un impulso anche se viene persa l'attività di membrana di una o più intercettazioni. E lo spessore dell'isolamento?
Rushton (1951) determinò che la velocità di propagazione del potenziale d'azione in una fibra nervosa sarà ottimale quando il rapporto tra il diametro interno della porzione mielinizzata della fibra e il suo diametro esterno è 0,6-0,7; questo indicatore è determinato dalla formula:
A vendita all'ingrosso =1/2e, dove K è il rapporto tra i diametri, e e= 2,718 . Misurazioni effettive di fibre nervose reali effettuate dagli istologi hanno confermato questi dati.
6. Classificazione delle fibre nervose in base alla velocità.
Sulla base degli studi elettrofisiologici dei nervi ottenuti da Erlanger e Gasser, tutte le fibre nervose sono divise in tre gruppi: A, B e C, con quattro sottogruppi identificati nel gruppo A (Tabella 1).
Un nervo può contenere un gran numero di fibre nervose, diverse nella velocità e nella direzione del movimento degli impulsi. Il nervo dell'arto inferiore nell'uomo, ad esempio, contiene fibre nervose che differiscono per funzione, diametro e velocità di conduzione degli impulsi. Il potenziale d'azione di un tale nervo è determinato da un intero spettro di gruppi di fibre e dalla velocità degli impulsi nervosi che li attraversano. Quando viene applicata un'irritazione che può eccitare le fibre nervose di tutti i gruppi, viene registrato un potenziale d'azione composito dell'intero nervo.
Tabella 1. Classificazione delle fibre nervose secondo Erlanger e Gasser].
|
Funzioni delle fibre |
Diametro medio della fibra, µm |
Velocità media di conduzione, m/s |
|
|
Fibre sensoriali e motorie dei muscoli scheletrici |
|||
|
Fibre afferenti del tatto e della pressione |
|||
|
Fusi muscolari motori |
|||
|
Dolore e temperatura sensibili alla pelle |
|||
|
Preganlionico simpatico |
|||
|
Postgangliare simpatico |
Il nervo dell'arto posteriore, ad esempio, contiene fibre nervose che differiscono per funzione, diametro e velocità di conduzione. Il potenziale d'azione di un tale nervo è determinato da un intero spettro di gruppi di fibre e velocità degli impulsi nervosi. Quando viene applicata un'irritazione che può eccitare le fibre nervose di tutti i gruppi, viene registrato un potenziale d'azione composito dell'intero nervo.
7. Generazione di eccitazione nei recettori.
In un organismo vivente, l'influenza degli stimoli naturali avviene in speciali organi di senso e gli stimoli sono luce, suono, pressione, temperatura, cambiamenti nel pH dell'ambiente, ecc. Le cellule che distinguono questi stimoli e inviano informazioni su di essi al sistema nervoso sono chiamate recettori.
Quando uno stimolo agisce su una cellula recettrice, si verifica la depolarizzazione della membrana dei suoi processi ricettivi e, con una forza dello stimolo sufficientemente grande, il cosiddetto potenziale del recettore. A differenza delle membrane di altre cellule eccitabili, la durata del potenziale recettore corrisponde alla durata dello stimolo e la sua ampiezza corrisponde all'intensità dello stimolo.
Il potenziale recettore si propaga elettrotonicamente alla base del corpo cellulare recettore, dove ha origine il suo assone, e qui provoca una depolarizzazione che supera il livello di soglia; si verifica un potenziale d'azione. Ciò che è specifico in questo caso è quanto segue: la curva di ripolarizzazione della membrana alla fine del potenziale d'azione scende molto al di sotto del livello iniziale, si verifica l'iperpolarizzazione della membrana e la successiva curva di depolarizzazione raggiunge nuovamente il livello di soglia: si verifica il potenziale d'azione successivo. Pertanto, una depolarizzazione persistente della membrana di ingresso di una cellula recettrice causata da uno stimolo provoca la generazione di una serie di potenziali d'azione sulla sua membrana di uscita, che poi si propagano lungo l'assone. La durata dell'emissione di serie di AP corrisponde alla durata dell'azione dello stimolo e la frequenza degli impulsi nella serie corrisponde all'intensità della sua azione. Pertanto, i potenziali d'azione sotto forma di un codice di frequenza trasmettono al sistema nervoso centrale l'entità e la durata dello stimolo.
I recettori che formano una singola unità con l'assone di una cellula nervosa sensibile sono detti primari; i recettori secondari sono collegati all'assone della cellula nervosa mediante sinapsi e la trasformazione del potenziale del recettore in una serie di potenziali d'azione avviene nelle terminazioni ricettive (terminali) della cellula nervosa sotto l'influenza del mediatore rilasciato dal recettore. La durata e la frequenza della serie in questo caso sono determinate dalla durata del rilascio e dalla quantità di mediatori.
8. Trasmissione intercellulare dell'eccitazione.
Le strutture che sorgono nel punto di contatto tra l'estremità dell'assone di una cellula nervosa e la membrana della cellula successiva (muscolare, ghiandolare o nervosa) sono chiamate sinapsi. La formazione chiamata sinapsi comprende membrane di cellule in contatto - presinaptiche e postsinaptiche, tra le quali rimane uno spazio largo 10-50 nm (100-500 A) - la fessura sinaptica. Questa ampiezza del divario rende impossibile il trasferimento elettrico dell'eccitazione da cellula a cellula a causa della grande perdita di corrente nel fluido intercellulare. La trasmissione chimica dell'eccitazione nelle sinapsi può tuttavia svolgere il ruolo di un meccanismo di rafforzamento dell'eccitazione. Dipende dalla quantità del mediatore rilasciato e dalla durata del suo rilascio, dalla sensibilità della membrana ricettiva. Tutto ciò può essere regolato in modo ottimale dall'evoluzione e, quindi, la sinapsi funziona secondo il principio di una valvola che consente l'eccitazione di un. una certa intensità da attraversare.
Il significato biologico delle sinapsi . Senza le sinapsi come valvole, l’attività ordinata del sistema nervoso centrale difficilmente sarebbe possibile. L'efficienza della trasmissione sinaptica può essere modificata. Ad esempio, la trasmissione dell'eccitazione migliora con un uso più frequente della sinapsi.
Il metodo sinaptico delle connessioni interneuronali gioca un ruolo importante nello sviluppo e nella formazione del sistema nervoso degli organismi nei processi di filogenesi e ontogenesi. Durante il processo di ontogenesi, il sistema nervoso centrale di un individuo si sviluppa e migliora fino ad una certa età, sia con l'aggiunta di nuovi neuroni, sia con la nascita e l'instaurazione di nuove connessioni sinaptiche interneuronali tra loro, cosa che sarebbe impossibile in caso di costante rigidità connessioni morfologiche intercellulari.
Le sinapsi, infine, sono il punto di applicazione di molte sostanze farmacologiche che bloccano la trasmissione dell'eccitazione, comprese sostanze di origine endogena che hanno significato farmacologico e patogenetico positivo. Saranno discussi quando si considererà la funzione delle strutture cerebrali.
Consideriamo il meccanismo della trasmissione sinaptica usando l'esempio della sinapsi neuromuscolare.
Giunzione neuromuscolare Con. La struttura della sinapsi neuromuscolare è mostrata schematicamente in Fig. 13.
Riso. 13. Sinapsi neuromuscolare (chimica).
Gli assoni dei motoneuroni del midollo spinale e i nuclei motori del midollo allungato e del mesencefalo formano sinapsi con le fibre muscolari scheletriche. La membrana presinaptica è formata da numerosi rami terminali dell'assone che hanno perso la guaina mielinica. La membrana postsinaptica della sinapsi appartiene alla fibra muscolare ed è chiamata placca terminale. Le vescicole sinaptiche nei terminali degli assoni ispessiti contengono una sostanza chimica chiamata acetilcolina.
Sotto l'influenza del potenziale d'azione dell'assone, l'acetilcolina viene rilasciata attraverso la sua membrana presinaptica dalle vescicole sinaptiche che, dopo aver attraversato la fessura sinaptica, provoca la depolarizzazione della placca terminale e la comparsa su di essa di un potenziale della placca terminale locale (EPP). Quest'ultimo differisce dal potenziale d'azione in numerose proprietà. Dura più a lungo; la sua curva, che si trova sul lato negativo dell'asse x, è caratterizzata da un forte aumento per 1-1,5 ms e un lento declino della durata di 3,5-6 ms. L'EPP non obbedisce alla legge "tutto o niente"; la sua ampiezza dipende dalla quantità di trasmettitore rilasciato nella sinapsi e dalla sensibilità della membrana postsinaptica ad esso. Con una frequenza sufficiente di potenziali d'azione che arrivano alla membrana presinaptica, la quantità di trasmettitore rilasciata nella sinapsi e, quindi, l'EPP si somma, stimolando la comparsa di un potenziale d'azione sulla membrana della fibra muscolare.
Per il normale rilascio del trasmettitore nella sinapsi è necessaria la presenza di ioni calcio. La ragione del rilascio del trasmettitore nella fessura sinaptica non è la depolarizzazione della membrana presinaptica stessa, ma il fatto che la depolarizzazione apre la strada al calcio nel terminale presinaptico. Se il calcio viene rimosso dall'ambiente esterno, la sinapsi chimica non funzionerà in caso di depolarizzazione. È in fase di studio il meccanismo d'azione degli ioni Ca 2+ sul rilascio del mediatore. È stato stabilito che il suo rilascio nella sinapsi è preceduto da un forte aumento della permeabilità della membrana presinaptica agli ioni calcio, che penetra rapidamente nel terminale presinaptico dell'assone. Dopo che gli ioni Ca 2+ hanno svolto la loro funzione, vengono pompati fuori dall'assone mediante pompe ioniche di calcio, il cui lavoro mantiene costantemente una concentrazione sbilanciata di ioni calcio attorno alla membrana.
Nei mitocondri dei terminali degli assoni viene prodotta l'energia necessaria al funzionamento delle pompe, che viene spesa anche per la risintesi dell'acetilcolina da colina e acido acetico negli stessi terminali.
Blocco della trasmissione neuromuscolare . Gli anestetici locali (novocaina) bloccano la conduzione dell'eccitazione nelle terminazioni nervose presinaptiche. La tossina botulinica rende impossibile il rilascio del trasmettitore poiché blocca la permeabilità della membrana presinaptica agli ioni calcio. Gli ioni magnesio agiscono in modo simile sulla membrana. Il veleno di serpente, ad esempio l'a-bungarotossina, il veleno di curaro, blocca i recettori dell'acetilcolina della membrana postsinaptica, in modo irreversibile (veleno di serpente) o reversibile (veleno di curaro) legandosi ad essi. I composti organofosforici in dosi tossiche inibiscono l'enzima colinesterasi, che normalmente scompone l'acetilcolina dopo aver svolto la sua funzione. In questo caso, l'enzima, che non viene scisso nel tempo, agisce nella sinapsi così a lungo che si verifica l'inattivazione dei recettori della membrana postsinaptica per il trasmettitore.
Dal veleno del ragno karakurt è stata isolata una proteina: la latrotossina, che è essenzialmente un canale del calcio che non chiude. È incorporato nella membrana presinaptica e inizia a far entrare il calcio nel terminale. Di conseguenza, le riserve di acetilcolina nei terminali sono completamente esaurite e il sistema nervoso non può causare contrazioni muscolari (comprese quelle respiratorie).
Alcune sostanze che bloccano la trasmissione sinaptica vengono utilizzate in anestesia e in altre procedure mediche per rilassare i muscoli. Queste sostanze sono chiamate rilassanti
Sinapsi elettriche . Oltre a quelle chimiche, nel corpo animale sono state trovate sinapsi elettriche. Nell'ES, i flussi di ioni passano dai terminali degli assoni alla cellula bersaglio direttamente attraverso canali speciali nei tubi di specifici dispositivi di “aggancio” delle proteine di membrana - connessoni. Le molecole proteiche della connessione formano una struttura speciale nelle membrane del terminale assonale e nella membrana postsinaptica della cellula bersaglio - connessione, composta da sei molecole e avente un canale all'interno. Due connettori di membrane sinaptiche adiacenti si collegano tra loro e in ciascuno di essi si apre un foro, un canale che era stato precedentemente chiuso (questo processo è simile all'apertura dei portelli durante l'attracco). Questo canale presenta una bassa resistenza al passaggio degli ioni. Ci sono molti connessioni in ES. Pertanto, l'ES collega due cellule con molti tubi sottili con un diametro di circa 1-1,5 nm, che passano all'interno delle molecole proteiche (Fig. 14, d).

Riso. 14. Sinapsi elettriche.
Lo spazio tra i connettori nella fessura sinaptica dell'ES è riempito con un isolante. Negli uccelli, nella catena di neuroni che assicurano la reazione della pupilla alla luce, lo spazio delle sinapsi elettriche è pieno di mielina.
Una caratteristica distintiva dell'ES è la sua velocità. La seconda caratteristica degli ES è che trasmettono il segnale in entrambe le direzioni, cioè sono simmetrici. Esistono, tuttavia, ES formati da membrane con proprietà rettificanti che trasmettono un segnale in una direzione, nonché sinapsi chimiche simmetriche, in cui sono presenti vescicole con un trasmettitore su entrambi i lati della fessura sinaptica. Gli ES si trovano più spesso negli invertebrati e nei vertebrati inferiori (ciclostomi e pesci).
Dispositivi come le sinapsi. Dispositivi come le sinapsi svolgono un ruolo importante nella vita di un'ampia varietà di tessuti e organi. Ad esempio, le cellule cardiache di diversi animali sono collegate da canali costituiti dalla stessa proteina connettiva che forma i canali nell'ES. In questo caso, l'area di contatto cellulare è chiamata contatto altamente permeabile (una sinapsi è un contatto in cui almeno un partecipante è un neurone). Le cellule muscolari lisce degli organi interni sono collegate tra loro da un sistema di connessioni. Anche le cellule non eccitabili (cellule epiteliali, ghiandole, fegato, ecc.) sono collegate da contatti altamente permeabili. Infatti, quasi tutti i tessuti del corpo non sono accumuli di singole cellule, ma un unico collettivo in cui le cellule possono scambiarsi varie molecole attraverso canali di contatto altamente permeabili. Grazie a ciò è possibile una sorta di “mutua assistenza cellulare” nei tessuti. Ad esempio, se le pompe a membrana non funzionano bene in una cellula, le cellule vicine, attraverso canali di contatti altamente permeabili, “condividono” con essa le loro riserve di ioni e ne mantengono il potenziale di riposo.
I connessoni non sono tubi stabili, ma strutture dinamiche: i canali connessoni possono aprirsi e chiudersi sotto l'influenza di vari fattori. Il meccanismo molecolare di questa chiusura del canale è stato chiarito. Connecton è costituito da 6 subunità che possono muoversi l'una rispetto all'altra (vedi Fig. 2-12, b) e il foro può chiudersi; Questo dispositivo è molto simile al dispositivo del diaframma di una fotocamera con lamelle mobili. Se una cellula ha subito un grave danno alla membrana e i suoi meccanismi protettivi non riescono a far fronte all'eccesso, ad esempio, di calcio proveniente dall'ambiente, a causa del quale la cellula è condannata a morte, l'elevata concentrazione di calcio all'interno di questa cellula influisce sulla i collegamenti con le cellule vicine, i loro canali si chiudono e le cellule vicine vengono disconnesse da quella vicina danneggiata. “Le cellule cardiache lavorano insieme ma muoiono da sole”, ha detto un famoso scienziato. Ora sappiamo perché questo è possibile.
Il dinamismo dei connessioni è importante non solo per la sopravvivenza. È stato accertato che contatti altamente permeabili si possono trovare già nelle prime fasi di sviluppo degli embrioni di vari animali; collegano le cellule che compaiono durante le prime divisioni dell'uovo e durante l'ulteriore sviluppo compaiono o scompaiono. Le cellule poi si influenzano a vicenda con alcune sostanze, poi le aree dell'embrione vengono isolate l'una dall'altra e in queste aree si sviluppa un tessuto omogeneo da cellule identiche; poi queste aree sono nuovamente collegate dai contatti con i vicini, e tutto questo complesso gioco di contatti svolge determinate funzioni nella regolazione dello sviluppo normale. Tutto ciò suggerisce che la segnalazione nell'ES è una “professione” secondaria della struttura che, come le pompe ioniche, svolge un ruolo più generale e fondamentale nello sviluppo degli organismi e nel funzionamento dei loro tessuti.
Allo stesso modo, il principio di funzionamento delle sinapsi chimiche viene utilizzato negli organismi non solo per trasmettere informazioni, ma anche per altri scopi. Pertanto, diverse cellule secretrici utilizzano gli ioni Ca2+ per regolare il rilascio delle secrezioni, proprio come in una sinapsi chimica questo processo viene utilizzato per rilasciare un trasmettitore. Si noti che le cellule di molte ghiandole sono elettricamente eccitabili.
Consideriamo, ad esempio, il lavoro delle cellule B del pancreas che producono insulina. Sulla superficie di queste cellule ci sono dei recettori che rispondono al livello di glucosio nel fluido tissutale che li lava. Se il livello di concentrazione di glucosio nel sangue è superiore al normale, sotto l'influenza di questi recettori le cellule vengono depolarizzate (a causa della chiusura dei canali del potassio) e in esse sorgono potenziali d'azione natura, si formano a causa dell'apertura dei canali del Ca. In questo caso, gli ioni Ca2+ entrano nella cellula, il che porta al rilascio di insulina nel sangue, come nel caso delle terminazioni nervose, porta al rilascio di un mediatore. Il ruolo del calcio nel rilascio di varie sostanze, in particolare degli ormoni, è dimostrato anche per molte altre ghiandole. È opportuno notare che il meccanismo descritto per la comparsa dei potenziali d'azione nelle cellule del pancreas probabilmente si verifica anche nelle cellule dell'ipotalamo, che rispondono a diversi livelli di concentrazione di alcuni ormoni,
Diverse cellule del corpo rilasciano diverse sostanze nel loro ambiente; Prima di tutto, queste sono cellule ghiandolari. Queste sostanze sono confezionate in contenitori a membrana - vescicole, e il loro rilascio è regolato da ioni Ca2+, che entrano nella cellula attraverso speciali canali del calcio. Come risultato dell'evoluzione, questo meccanismo cominciò ad essere utilizzato dalle cellule nervose nella costruzione delle sinapsi chimiche; i contenitori contengono un mediatore, quindi il suo rilascio è organizzato allo stesso modo del rilascio di ormoni e altre sostanze, e da questo punto di vista le cellule nervose con sinapsi chimiche sono una delle varianti delle cellule secretrici, e il mediatore è il loro segreto, che non solo viene rilasciato nel sangue e raggiunge consumatori molto specifici attraverso la fessura sinaptica. D’altra parte, il lavoro delle cellule ghiandolari è simile al lavoro delle cellule nervose, ma il loro potenziale d’azione serve ad aprire i canali del calcio e a far entrare gli ioni Ca2+. Osserviamo la stessa cosa nelle cellule muscolari, dove il potenziale d'azione apre la porta agli ioni Ca 2+, che innescano il processo di contrazione.
Pertanto, in primo luogo, vediamo ancora una volta la stretta somiglianza dei meccanismi utilizzati dalle diverse cellule del corpo per scopi diversi, e in secondo luogo, guardando al futuro, possiamo suggerire che il segreto della colorazione emotiva delle sensazioni e della più alta manifestazione del sistema nervoso l'attività del cervello umano sono le sensazioni della coscienza e del pensiero - si trovano proprio in questa unità di funzioni neuronali. quelli. ha la proprietà di essere sia una cellula nervosa (generatrice e conduttrice di impulsi elettrici) che una cellula secernente.
9. Dipendenza della funzione di un neurone dalla sua forma.
Le funzioni fisiologiche di una cellula nervosa sono in gran parte determinate dalle sue proprietà elettriche. Sappiamo già che i neuroni hanno le forme più diverse, con gli assoni generalmente a forma di cavo cilindrico e il corpo più o meno sferico. Confrontando le proprietà elettriche di una sfera e di un cilindro formato da membrane identiche, vedremo quanto sia importante la forma della cellula eccitabile nel determinare queste proprietà.
I parametri elettrici che caratterizzano una cellula nervosa possono essere suddivisi in due gruppi: 1) parametri caratterizzanti la sostanza della membrana e del citoplasma - la resistenza specifica della membrana (Rm), pari a 1-10 kOhm/cm2, la sua capacità elettrica specifica (1 μF/cm2) e resistenza citoplasmatica specifica (Ri) pari a circa 100 Ohm.cm; questi parametri non dipendono dalla forma della cella; 2) parametri di sistema; caratterizzano la fibra, la cellula o il sistema di cellule connesse nel loro insieme e dipendono dalla dimensione e dalla forma; uno dei parametri del sistema è la resistenza di ingresso - Rin
La resistenza di ingresso è il rapporto tra lo spostamento del potenziale di membrana della cellula e la quantità di corrente elettrica passata al suo interno: Rin = DV/I. Pertanto, la resistenza di ingresso è un analogo della normale resistenza di una sezione di fibra nervosa.
La resistenza di ingresso dell'assone (cilindro) è espressa dalla formula:
Rin = 1/p2(Rm Ri)/8a3, Dove UN- raggio dell'assone .
In questa formula, Rm e Ri sono rispettivamente la resistenza della membrana e del citoplasma dell'assone, cioè parametri del 1° gruppo; tutto il resto è determinato dalla geometria della fibra come un cilindro la cui lunghezza è molto maggiore del suo raggio. Cosa succederebbe se una cella sferica di raggio a fosse fatta dello stesso materiale? È stato dimostrato che anche in cellule sferiche molto grandi (1 mm di diametro), la resistenza citoplasmatica è solo lo 0,1% della resistenza di membrana; nelle cellule sferiche di diametro significativamente più piccolo (tutte le cellule nervose sono così), la resistenza del citoplasma può essere ignorata, e in realtà la resistenza di ingresso di una cellula sferica dipende solo dalla resistività della sua membrana e del suo raggio: Rin = Rm / 4pa3
Pertanto, per una cella sferica, la resistenza di ingresso è direttamente proporzionale alla resistenza della membrana, quindi se, quando la cella è eccitata, la resistività della sua membrana diminuisce, diciamo, di 36 volte, allora anche la resistenza di ingresso del sistema diminuirà per 36 volte.
Per una fibra, la resistenza di ingresso è proporzionale solo a 2Rm, quindi se durante l'eccitazione anche la resistenza di membrana diminuisce di 36 volte, allora la resistenza di ingresso cambierà solo di 6 volte.
Come incide tutto ciò sulle funzioni del neurone? In una cellula sferica, il potenziale di membrana non scompare istantaneamente, come in una fibra cilindrica, ma lentamente, in modo esponenziale. Il potenziale postsinaptico nelle sinapsi è direttamente proporzionale alla resistenza di ingresso della membrana postsinaptica, quindi in una cellula sferica di diametro. di 10 micron, il potenziale postsinaptico sarà 100 volte maggiore che in una cellula con un diametro di 100 µm. Minore è la resistenza di ingresso, minore è l'efficienza delle sinapsi eccitatorie. Il mediatore della sinapsi inibitoria nella cellula globulare riduce la resistenza all'ingresso della membrana postsinaptica, riducendo così l'efficacia degli impulsi eccitatori; nella fibra cilindrica questo effetto è molto minore.
Una lenta diminuzione del potenziale di membrana sulla membrana di una cellula sferica aumenta significativamente l'efficienza della somma temporanea in essa. Le diverse forme e dimensioni dei corpi dei neuroni nella corteccia cerebrale non sono un fenomeno casuale!
Conduttività di una fibra nervosa al variare del suo diametro. È stato dimostrato che nella stessa fibra, prima della sua espansione, l'impulso nervoso rallenta e la sua ampiezza diminuisce; Quando la fibra si espande sei volte, il PD non può passare attraverso questa espansione. Al contrario, quando la fibra si restringe, man mano che si avvicina al sito del restringimento, la velocità dell'impulso aumenta e la sua ampiezza aumenta. Un aumento dell'ampiezza AP all'estremità del terminale è molto importante per il funzionamento delle sinapsi chimiche, poiché migliora le condizioni per il rilascio dei mediatori Se un assone affusolato si ramifica in due rami, il cui diametro totale è inferiore a il diametro nel sito di ramificazione, quindi l'AP prima della ramificazione accelera e, se il diametro totale dei rami è maggiore, la velocità PD rallenta e potrebbe persino bloccarsi. Conduttività dei dendriti. Nella direzione della conduzione dell'eccitazione, i dendriti iniziano con rami sottili e numerosi, che si fondono ripetutamente in rami più spessi e terminano con tronchi spessi sul corpo del neurone. La lunghezza dei dendriti è decine di volte maggiore del diametro del corpo delle cellule nervose e lo spessore dei rami terminali può essere di frazioni di micrometro. In alcuni dendriti la membrana non è eccitabile (non genera AP) e può trasmettere segnali solo elettrotonicamente come un cavo passivo; in altri è capace di generare e condurre potenziali d'azione come un assone, ma in direzione del corpo del neurone; Nel primo caso, secondo gli scienziati, molte sinapsi si trovano su sottili rami terminali e l'azione congiunta di queste sinapsi raggiunge il corpo del neurone. Una valutazione quantitativa ha mostrato che l'efficienza delle sinapsi dendritiche è solo 3-5 volte inferiore a quella delle sinapsi situate sul corpo del neurone. Il potenziale elettrotonico viene ricevuto nella sinapsi dendritica a causa del piccolo raggio del terminale dendritico e. quindi le sue grandi dimensioni per raggiungere efficacemente la resistenza di ingresso del corpo cellulare (vedi sopra). Questa forza è sufficiente a compensare l'attenuazione del segnale mentre si muove nella direzione dell'espansione dendritica.
Nei neuroni la cui membrana dendritica è in grado di generare e condurre potenziali d'azione, l'elevata efficienza simile delle sinapsi dendritiche porta al fatto che solo poche sinapsi porteranno il potenziale di membrana alla soglia e causeranno un potenziale d'azione in questo ramo che inizierà a diffondersi nel corpo cellulare. Il suo ulteriore destino dipenderà dalle proprietà dei nodi di ramificazione che dovrà attraversare nel percorso verso il corpo del neurone, dal numero e dalla coincidenza temporale dei potenziali d'azione che arrivano ai siti di ramificazione lungo altri rami, ecc. Di conseguenza, questo tipo di cella funziona come un circuito logico complesso. Le cellule con un complesso sistema di dendriti funzionano come elementi informatici molto complessi.
Funzioni delle spine dendritiche . Sui dendriti di molti neuroni ci sono formazioni speciali, le cosiddette. spine. Queste formazioni simili a funghi, a forma di testa su un sottile gambo, rappresentano una sporgenza della membrana cellulare di un dendrite su cui si forma una sinapsi chimica con il terminale di un altro neurone. Gli scienziati ritengono che la funzione delle spine sia la seguente. Nel caso di una membrana non eccitabile, le spine riducono significativamente l'influenza reciproca delle sinapsi vicine sulla membrana postsinaptica del dendrite, nella quale, in assenza di spine, si avrebbe una significativa diminuzione della resistenza di ingresso del dendrite terminale e, di conseguenza, una diminuzione del livello del potenziale di membrana. I calcoli hanno dimostrato che, sebbene le sinapsi della colonna vertebrale siano singolarmente meno efficaci delle sinapsi situate direttamente sul dendrite, l'effetto totale del loro lavoro è notevolmente maggiore.
Se la membrana della colonna vertebrale è eccitabile, può funzionare come amplificatore della trasmissione sinaptica. A causa della sottigliezza del collo, la resistenza di ingresso della colonna vertebrale è molto elevata e una sinapsi può causare un potenziale d'azione nella testa, che invierà al dendrite una corrente elettrica molto più forte della corrente della sinapsi.
10. Funzioni della neuroglia.
Lo spazio tra i neuroni nel sistema nervoso centrale è pieno di cellule di supporto specializzate chiamate glia. Il loro numero è 5-10 volte superiore a quello dei neuroni. Glia formano anche le guaine mieliniche degli assoni periferici dei neuroni.
Le cellule gliali sono divise in due gruppi: astrociti e oligodendrociti. Gli astrociti puliscono lo spazio extracellulare dei neuroni dagli ioni e dai mediatori in eccesso e quindi agiscono come un tampone in caso di aumento della concentrazione extracellulare di ioni calcio ed eliminano le "interferenze" chimiche durante. il funzionamento dei neuroni. La funzione trofica delle glia in relazione ai neuroni non è stata dimostrata, sebbene possano alterare l'accesso di ossigeno e glucosio ai neuroni. bloccando i corrispondenti flussi di liquido intercellulare. Gli astrociti possono modificare il segnale trasmesso alle sinapsi bloccando alcune sezioni delle membrane nelle fessure sinaptiche. In luoghi con danni cerebrali limitati, gli astrociti svolgono funzioni sanitarie, partecipando alla rimozione di pezzi morti di neuroni.
Gli oligodendrociti avvolgono gli assoni dei neuroni e formano la loro guaina mielinica nel sistema nervoso centrale. Un tipo di oligodendrociti, le cellule di Schwann, formano la guaina mielinica discontinua degli assoni periferici.
11.Crescita e sviluppo di un neurone nell'ontogenesi.
Un neurone si sviluppa da una piccola cellula precursore che smette di dividersi ancor prima di produrre i suoi processi.
La questione della divisione dei neuroni stessi rimane attualmente controversa. così come l'opportunità stessa di questo processo nell'organismo formato. La rigenerazione dei processi danneggiati dal corpo cellulare è un fatto pienamente dimostrato. In genere, l'assone inizia a crescere per primo e i dendriti si formano successivamente. Alla fine del processo di sviluppo della cellula nervosa appare un ispessimento di forma irregolare - un "cono di crescita", che, apparentemente, si fa strada attraverso il tessuto circostante. Il cono di crescita è una parte appiattita del processo delle cellule nervose con molte spine sottili. Le micropunte hanno uno spessore di 0,1-0,2 µm e una lunghezza massima di 50 µm. Per fare un confronto, ricorda che il diametro di un globulo rosso umano è di 7,3 micron. La regione ampia e piatta del cono di crescita è di circa 5 µm in larghezza e lunghezza. Gli spazi tra le microspine sono ricoperti da una membrana ripiegata.
I microspighe sono in costante movimento. Alcuni di essi si ritraggono nel cono, altri si allungano, deviano in direzioni diverse, toccano il substrato e possono attaccarsi ad esso. Direttamente sotto le aree piegate della membrana e nelle spine si trova una massa densa di filamenti di actina, mitocondri, microtubuli e neurofilamenti aggrovigliati simili a quelli del corpo del neurone.
È probabile che i microtubuli, i neurofilamenti e i filamenti di actina (vedi sotto) si allunghino principalmente a causa dell'aggiunta di subunità appena sintetizzate alla base del processo neuronale. Si muovono ad una velocità di circa un millimetro al giorno, che corrisponde alla velocità del lento trasporto assonale in un neurone maturo. Viene aggiunto il nuovo materiale della membrana, apparentemente alla fine. Piccole vescicole di membrana vengono trasportate lungo il processo neuronale dal corpo cellulare al cono di crescita con un flusso di trasporto assonale veloce. Il materiale della membrana viene apparentemente sintetizzato nel corpo del neurone, trasportato al cono di crescita sotto forma di vescicole e qui incorporato nella membrana plasmatica per esocitosi, allungando così il processo della cellula nervosa.
La crescita degli assoni e dei dendriti è solitamente preceduta da una fase di migrazione neuronale, quando i neuroni immaturi si disperdono e trovano una sede permanente nelle reti nervose.
Il potenziale d'azione si muove lungo l'assone a causa della propagazione longitudinale della corrente. Ciascuna sezione della membrana, generando un impulso secondo il principio “tutto o niente”, eccita la sezione vicina con la sua depolarizzazione. Per comprendere il meccanismo di propagazione dell'impulso, nonché la trasmissione e l'integrazione sinaptica, è necessario comprendere il principio della propagazione passiva dell'impulso lungo il nervo.
Quando la corrente si muove lungo l'assone o il dendrite, perde la sua forza. La diminuzione della corrente dipende da diversi motivi, principalmente dal diametro e dalle proprietà della membrana della fibra nervosa. Maggiore è il diametro della fibra e maggiore è la resistenza della membrana, maggiore è la propagazione longitudinale della corrente. Le proprietà capacitive della membrana influenzano l'andamento temporale dei segnali elettrici, nonché la propagazione della corrente. Per stimare la distanza alla quale può propagarsi un potenziale sottosoglia, è necessario conoscere la geometria e le proprietà della membrana neuronale e, inoltre, l'andamento temporale dei potenziali cambiamenti.
Gli assoni di molti tipi di cellule nervose nei vertebrati sono ricoperti da una guaina mielinica, che ha un'elevata resistenza e una bassa capacità. La guaina mielinica funge da bobina isolante del filo e fa sì che la corrente si muova lungo la membrana. In questo caso, la corrente salta rapidamente da un nodo di Ranvier (un breve tratto di membrana privo di mielina) all'altro e la velocità di conduzione aumenta. Le fibre mielinizzate si trovano in aree della rete neurale dove la velocità di conduzione gioca un ruolo importante.
I segnali elettrici possono anche essere trasmessi da un neurone all'altro in punti di stretto contatto tra loro, chiamati giunzioni comunicanti. La corrente in tali connessioni scorre attraverso canali speciali, connessioni.
Proprietà elettriche passive delle membrane nervose e muscolariLa permeabilità delle membrane neuronali e il modo in cui contribuiscono alla generazione di potenziali d'azione sono stati discussi nei capitoli precedenti. Questo capitolo si concentrerà principalmente su come le correnti si propagano lungo una fibra nervosa e creano differenze di potenziale locale.
Le proprietà elettriche passive dei neuroni, vale a dire la resistenza e la capacità di membrana, nonché la resistenza citoplasmatica, svolgono un ruolo critico nella segnalazione neuronale. Negli organi di senso queste proprietà costituiscono il legame tra uno stimolo sensoriale e la generazione di un impulso; a livello dell'assone permettono la propagazione dell'impulso; a livello sinaptico determinano la capacità del neurone postsinaptico di aggiungere e sottrarre potenziali sinaptici derivanti da numerosi input sinaptici, sia in prossimità del corpo cellulare che sui dendriti più distanti. Per comprendere questi processi è necessario conoscere i principi di propagazione dei segnali elettrici lungo i processi neurali. Questo capitolo si concentrerà principalmente sulle fibre nervose con diametro costante per tutta la loro lunghezza, cioè conduttori cilindrici. Inoltre, per questa descrizione assumeremo che in assenza di rigenerazione la membrana sia veramente passiva, vale a dire che le variazioni di potenziale che non raggiungono un livello di soglia non attivano conduttanze voltaggio-dipendenti e quindi non modificano la resistenza della membrana. Questi principi si applicano anche a strutture più complesse, come terminali assonici ramificati o arborizzazioni dendritiche con proprietà elettriche eterogenee. Il ruolo di tali strutture nel funzionamento del sistema nervoso è molto importante, ma una descrizione quantitativa delle loro proprietà elettriche richiederebbe un'analisi più complessa.
Proprietà del cavo delle fibre nervose e muscolari
Una fibra nervosa cilindrica è costituita dagli stessi componenti di un cavo elettrico sottomarino: un'asta conduttrice e una guaina isolante avvolta da un mezzo conduttivo. Tuttavia, la differenza quantitativa tra questi due sistemi è molto ampia. L'anima del cavo è solitamente in rame o un metallo ad altissima conduttività, mentre la guaina è in plastica o altri materiali ad altissima resistenza. Inoltre, l'involucro è solitamente piuttosto spesso e quindi ha una capacità ridotta. La tensione applicata a tale filo può essere trasmessa su una distanza considerevole grazie al fatto che la resistenza del rame è bassa, così come le perdite attraverso la guaina. Il contenuto della fibra nervosa è una soluzione di sali, simile in concentrazione al mezzo extracellulare e, a differenza del rame, ha una scarsa conduttività. La membrana cellulare, a sua volta, non è un buon isolante e ha un'elevata capacità a causa del suo piccolo spessore. La tensione applicata a una fibra nervosa non percorre una distanza significativa per due motivi:
1) la conduttività del contenuto di fibre è bassa, quindi la resistenza attuale è elevata;
2) la corrente che scorre lungo il citoplasma viene dissipata a causa di perdite attraverso la membrana, che non fornisce un isolamento sufficiente.
L'analisi della corrente via cavo fu iniziata da Lord Kelvin in relazione alle comunicazioni telefoniche transatlantiche e migliorata da Oliver Heaviside. Alla fine del 20 ° secolo. Heaviside fu il primo a tenere conto dell'importanza della corrente dispersa attraverso una guaina isolante, equivalente ad una membrana cellulare, e apportò anche molte importanti aggiunte alla teoria dei cavi, inclusa la definizione del concetto di impedenza. La teoria del cavo fu utilizzata per la prima volta per le fibre nervose da Hodgkin e Rushton, che misurarono sperimentalmente la propagazione del potenziale d'azione in un assone di aragosta utilizzando elettrodi extracellulari. Successivamente, elettrodi intracellulari furono utilizzati per misurazioni simili in un certo numero di fibre nervose e muscolari.
La regola di base qui è la legge di Ohm: la corrente i, passando attraverso la resistenza r, crea una tensione ν = ir. Di seguito considereremo anche l'influenza della capacità di membrana sull'entità e sull'andamento temporale della propagazione longitudinale della corrente.
Corrente del cavo
Per comprendere meglio i principi del flusso di corrente attraverso un cavo, immaginiamo come il calore si diffonde lungo un'asta metallica inserita in una guaina termoisolante, posta in un mezzo conduttore (ad esempio acqua). Se un'asta viene riscaldata da un'estremità, il calore si diffonde lungo l'asta e, diffondendosi, viene parzialmente dissipato nell'ambiente penetrando attraverso la guaina. Più ci si allontana dall'estremità riscaldata, più bassa è la temperatura; Quando la temperatura diminuisce, la velocità di dissipazione del calore diminuisce. A condizione che l’ambiente abbia un’elevata conduttività termica, la distanza su cui si diffonderà il calore dipenderà principalmente da due parametri:
1) dalla conduttività termica dell'asta
2) sulle proprietà isolanti del guscio.
Il flusso di corrente in un cavo avviene in modo simile. Una tensione applicata a un'estremità del cavo fa sì che la corrente fluisca lungo l'asta in direzione longitudinale, che viene parzialmente dispersa attraverso la guaina nell'ambiente. Quanto più è lontana l'estremità a cui viene applicata la tensione, tanto minore è la corrente. La distanza percorsa dalla corrente dipenderà dalla conduttività dell'asta e dall'efficacia con cui la guaina previene la perdita di corrente. Il guscio a bassa resistenza consentirà a tutta la corrente di passare nell'ambiente. Una guaina con una resistenza maggiore permetterà alla corrente di percorrere una distanza maggiore.
La corrente in un assone è trasportata dagli ioni: quando la corrente viene iniettata in una fibra nervosa (come un assone di aragosta) attraverso un microelettrodo, le cariche positive iniettate respingeranno altri cationi e attireranno gli anioni. Il più abbondante dei piccoli ioni all'interno della cellula è il potassio, che quindi trasporta la maggiore quantità di corrente attraverso la membrana. La corrente scorre longitudinalmente lungo l'assone e, mentre si muove, parte di essa viene persa a causa del movimento degli ioni attraverso la membrana. In una membrana con bassa resistenza e alta conduttività ionica, la maggior parte della corrente andrà persa prima che abbia il tempo di percorrere una distanza significativa. Con una maggiore resistenza della membrana, la corrente percorrerà una distanza maggiore lungo l’assone prima di dissiparsi nell’ambiente.
Post correlati:
 "Grazie al compagno Stalin per la nostra infanzia felice" Grazie al compagno Stalin per la nostra infanzia felice
"Grazie al compagno Stalin per la nostra infanzia felice" Grazie al compagno Stalin per la nostra infanzia felice
 Zhirinovsky preme, i comunisti restano indietro
Zhirinovsky preme, i comunisti restano indietro
 Storia dei Rurikidi e del loro albero genealogico
Storia dei Rurikidi e del loro albero genealogico
 Tolling materie prime: contabilità Schema di tolling 1c
Tolling materie prime: contabilità Schema di tolling 1c
 È possibile falciare il trifoglio strisciante?
È possibile falciare il trifoglio strisciante?





